总论

{kind=link}
图1:有丝分裂基本过程

{kind=link}
图2:小鼠的成纤维细胞分裂时会蜷缩成一团

{kind=link}
图3:出芽酵母的芽的大小对应着其在细胞周期中的位置


{kind=link}
图4:用人造核苷酸标记正在复制DNA的细胞


{kind=link}
图5:用流式细胞仪检测每个细胞中DNA含量
- 所有真核生物都通过有丝分裂(Mitosis)和减数分裂(Meiosis)复制。(本文只讨论有丝分裂)
- 大多数细胞在有丝分裂时,核膜解体而细胞核外的中心体牵引染色体。
- 有些低等生物的细胞有丝分裂时,核膜不解体,中心体移至细胞核内牵引染色体,早期被误认为没有中心体,故称“无丝分裂”(Amitosis),此词现已废止。
- 高等生物的细胞发生细胞凋亡时,会破裂为许多碎片,称为凋亡小体,早期也曾被误认为一种“无丝分裂”。
- 草履虫行无性生殖时,只有小核DNA复制且有中心体牵引;大核只能随机将染色体分散至两个子细胞,可能是真核生物中唯一的“无丝分裂”。
- 从一次有丝分裂结束到下一次有丝分裂结束称为一个细胞周期(Cell Cycle)。
- 有丝分裂由S期和M期组成,S期发生DNA复制,M期发生细胞分裂。
- 上一次有丝分裂结束到下一次S期开始,之间的间隔称为G1期。
- 从S期结束到M期开始,之间的间隔称为G2期。
- S期很长,可达10-12 h;M期很短,只需不到1 h。
- 有些细胞需要等待很长时间甚至永远不进入S期,这些细胞称为处在G0期。
- 有丝分裂基本过程见图1。
- 常用的研究有丝分裂的模式物种:出芽酵母(Saccharomyces Cerevisiae)、裂殖酵母(Schizosaccharomyces Pombe)、爪蟾(Xenopus Laevis)、果蝇(Drosophila Melanogaster)。
- 观察有丝分裂的方法:
- 小鼠的成纤维细胞在有丝分裂时会蜷缩成一团。(见图2左下角)
- 观察出芽酵母的芽的大小可判断其在细胞周期中的位置。(图3)
- 用荧光染料给DNA染色;或用抗体标记微管蛋白,可判断细胞在细胞周期中的位置。
- 在培养基中加入溴脱氧尿苷(Bromodeoxyuridine,一种人造核苷酸),可标记正在复制DNA的细胞。(图4)
- 计算处在S期的细胞的比例,可知S期在细胞周期中大约比例。
- 用流式细胞仪(Flow Cytometer)可检测每个细胞中DNA含量,从而知道处在S期和M期的细胞的比例。(图5)
- 处在M期的细胞的比例,称为有丝分裂指数(Mitotic Index)。
细胞周期控制系统


{kind=link}
图6:细胞周期控制系统

{kind=link}
图7:常见Cdk和周期素


{kind=link}
图8:各周期素在细胞周期中的浓度变化

{kind=link}
图9:Cdk的激活

{kind=link}
图10:APC/C和SCF酶的激活


{kind=link}
图11:细胞周期调控蛋白的其它信息
- 动物的早期胚胎细胞中,细胞周期的每个过程都有固定时间,不会因为某个事件未完成而暂停。
- 大多数细胞都有一套复杂的信号通路,控制细胞周期各阶段间的转换。
- 细胞周期控制系统的核心是两类蛋白:周期素(Cyclin,又名细胞周期蛋白)和依赖周期素的激酶(Cyclin-dependent Kinase,下文简称Cdk)。
- 细胞周期的控制有三个关键点:
- 从G1期到S期,称为Start,脊椎动物中又名限制点(Restriction Point)。
- 从G2期到M期。
- 从中期到后期。
- Cdk与特定的周期素结合时,能磷酸化特定的蛋白质,从而推动细胞周期前进。
- 不同的周期素在细胞周期的不同阶段被合成或降解,其浓度有规律地起伏,故称“周期素”。
- 酵母细胞只有一种Cdk,它与不同周期素结合时变为不同构象。
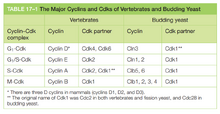
- 脊椎动物有四种Cdk,分别记为Cdk1、Cdk2、Cdk4、Cdk6。
- 常见周期素有四类:
- G1/S周期素在G1期上升,促使细胞进入S期,在S期下降。
- G1周期素帮助调控G1/S周期素,在G1期上升,在M期下降。(有些细胞没有此周期素)
- S周期素在G1期上升,促进染色体复制,在M期下降。
- M周期素在S期上升,促进细胞进入M期,在M期下降。
- 周期素不仅有激活Cdk的功能,也有把它引导至特定蛋白质的功能。
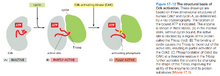
- Cdk的完全激活不仅需要周期素,还需在结合周期素后被Cdk激酶(Cdk-activating Kinase,CAK)磷酸化。
- 若Cdk作用位点的顶部的一对氨基酸残基被Wee1酶磷酸化,则会抑制Cdk活性;Cdc25酶则能将它们脱磷酸化,促进Cdk活性。
- 若Cdk结合周期素后结合Cdk抑制蛋白(Cdk Inhibitor Protein,CKI),则会抑制Cdk活性。(p27蛋白是一种CKI)
- CKI主要被用于控制G1/S和S周期素。
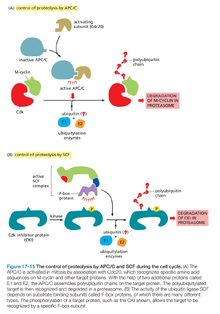
- 从中期到后期的转换主要不通过蛋白质磷酸化完成,而是通过周期体(Cyclosome,又名后期促进复合体,Anaphase-promoting Complex,或APC/C)将特定蛋白用泛素标记降解完成。
- APC/C降解的主要蛋白有:Securin(导致姐妹染色单体分离,见下文)、S周期素、M周期素。(但没有G1周期素)
- APC/C通过与Cdc20蛋白(有丝分裂中期)或Cdh1蛋白(有丝分裂后期至G1期)结合而激活。
- APC/C在G1/S周期素上升时被关闭。
- 细胞周期中的另一个泛素标记酶是SCF,用于在G1末期降解一些CKI蛋白,和在S期降解G1/S周期素。
- SCF通过与F-box蛋白结合而激活。
- 关于细胞周期调控蛋白的其它信息见图11。
S期


{kind=link}
图12:DNA复制的调控


{kind=link}
图13:黏连蛋白可能是把两条染色单体绑在一起的


{kind=link}
图14:DNA甲基化的三种状态
注:本文不讨论DNA复制的具体过程,它被留给另一篇文章。
- 染色体的复制起始点处,在细胞周期全程都有起始点识别复合体(Origin Recognition Complex,ORC)结合。
- M期结尾至G1期开头,Cdc6、Cdt1蛋白和ORC将无活性的DNA解旋酶装至起始点附近,与ORC组成预复制复合体(Prereplicative Complex,PreRC)。
- S-Cdk磷酸化特定蛋白,引起DNA解旋酶激活和DNA复制复合体装配。
- DDK(另一种激酶)磷酸化DNA解旋酶,亦促进DNA复制。
- 细胞防止DNA复制两次的措施:
- S-Cdk磷酸化ORC和Cdc6蛋白,阻止新DNA复制复合体产生。
- APC/C在G1期被关闭,防止它降解联会蛋白(Geminin),而联会蛋白抑制Cdt1蛋白。
- Cdt1蛋白与复制叉处的一个蛋白结合时会促进其降解。
- S-Cdk能促进核小体的4种组蛋白的合成,为染色质结构复制提供原料。
- 组蛋白被复制叉处的核小体组装因子分配到新复制出的染色质上。
- 染色质的表观修饰如何被复制尚未研究清楚。
- DNA甲基化研究得较为清楚:
- 哺乳动物有三种DNA甲基转移酶(DNA Methyltransferase):DNMT1、DNMT3A、DNMT3B。(DNMT2结构上与这几种酶类似,但它是tRNA甲基转移酶;DNMT3L参与DNA甲基化的调控,但它没有酶活性)
- 若DNA的两条链都有甲基化修饰,称为完全甲基化的(Fully Methylated)。
- 刚完成复制的DNA,其中一条链无甲基化修饰,称为半甲基化的(Hemimethylated)。
- 若半甲基化的DNA被复制,则其中一条子DNA两条链都无甲基化修饰,称为非甲基化的(Ummethylated)。
- 将非甲基化的DNA变为完全甲基化,需要从头甲基化酶(de novo Methyltransferase),一般由DNMT1完成。
- 将半甲基化的DNA变为完全甲基化,需要维护甲基化酶(Maintenance Methyltransferase),一般由DNMT3A和DNMT3B完成。(但这两类酶也各能以较低活性完成另一类的工作)
- DNMT3A主要修饰卫星片段,DNMT3B主要修饰常染色质。
- 缺少DNMT3A的小鼠能出生,但不能产生生殖细胞,且患有无神经节性巨肠节(Aganglionic Megacolon)。
- 缺少DNMT3B的小鼠在胚胎晚期死亡。
- DNMT3A和DNMT3B同时缺少,或缺少DNMT1的小鼠在胚胎早期即死亡。
- 组蛋白修饰如何复制研究得不清楚:
- 原有的有修饰的组蛋白在DNA复制时被暂时取下,复制后每个会被随机装到两条子DNA的一条上,且另一条的对应位置一般是无修饰的组蛋白。
- 目前鉴定出了一些酶,会在DNA复制结束时修饰组蛋白,但尚不知它们如何获取模板,另外不是所有组蛋白修饰的酶都被发现。
- 有种理论认为,组蛋白修饰不是完全在DNA复制时进行的,而是部分在基音转录时进行。
- DNA甲基化研究得较为清楚:
- 复制出的两条染色体通过黏连蛋白(Cohesin)结合在一起。(此时两条染色体各称为染色单体,结合在一起组成的结构称为一条染色体)
- 黏连蛋白具有环状结构,可能是通过这个环把染色单体绑在一起。(图13)
- 染色单体结合的另一方式是通过DNA缠绕(Catenation)。
- 在M期,拓扑异构酶II将缠绕的两条DNA链解开。
- S期中心粒会复制,由G1/S-Cdk启动,但两个中心粒仍被包在一个中心体中。
M期


{kind=link}
图15:M-Cdk的激活

{kind=link}
图16:前期核膜的变化


{kind=link}
图17:凝缩蛋白


{kind=link}
图18:三种纺锤丝


{kind=link}
图19:四种马达分子


{kind=link}
图20:染色体经多次尝试才能正确地与纺锤丝相连
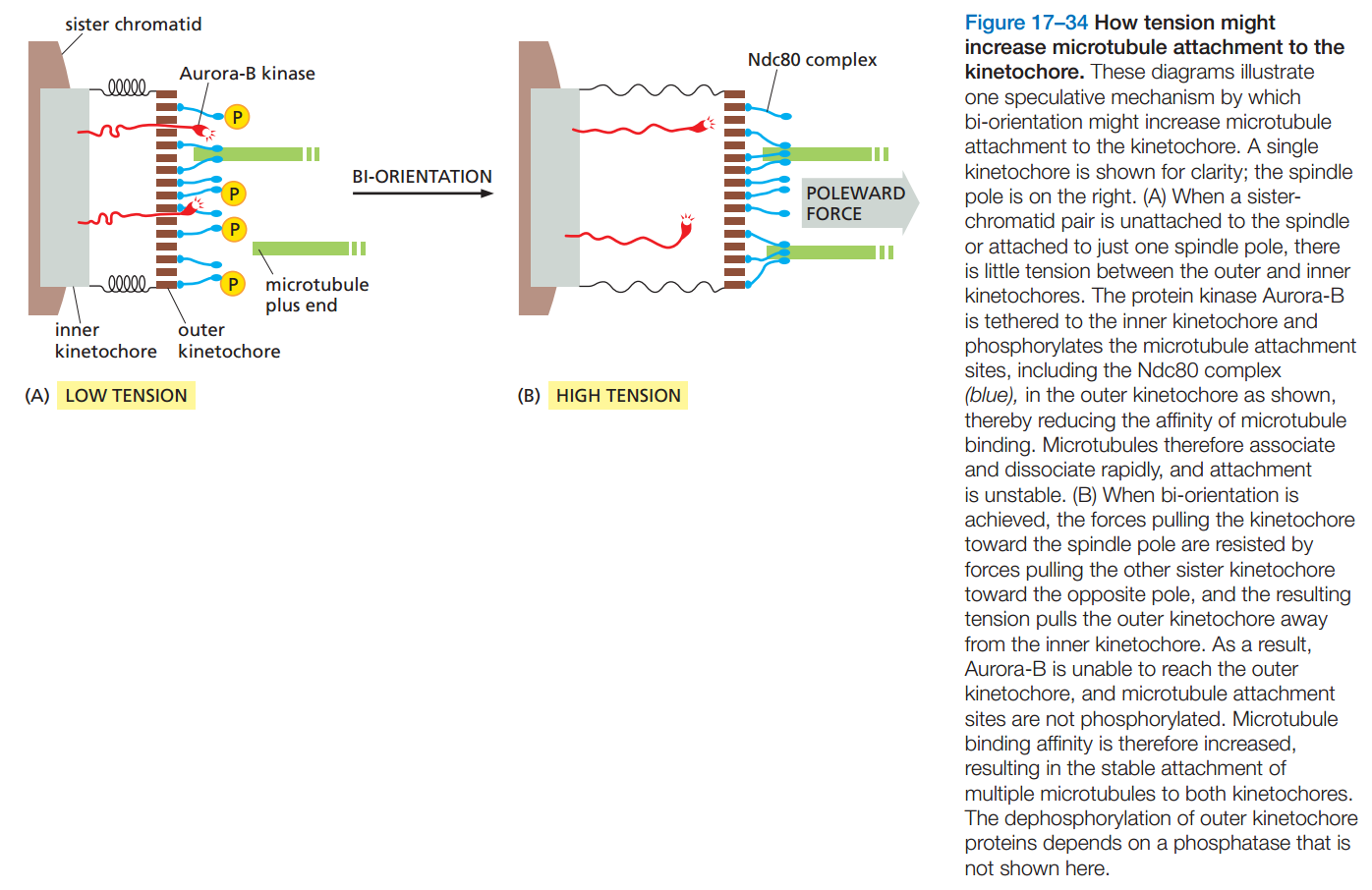

{kind=link}
图21:张力是保持正确连接稳定的原因


{kind=link}
图22:从中期到后期


{kind=link}
图23:染色体向中心体移动时,微管正极解体


{kind=link}
图24:中体


{kind=link}
图25:收缩环的启动


{kind=link}
图26:收缩环位置确定的三种理论
- M期有两个主要事件:核分裂(Mitosis)、胞质分裂(Cytokinesis)。
- M期可分为两部分:由M-Cdk主导的部分、由APC/C主导的部分。
- M期在形态上可分为五部分:前期(Prophase)、前中期(Prometaphase)、中期(Metaphase)、后期(Anaphase)、末期(Telophase)。
- 核分裂贯穿M期全程,胞质分裂只在末期进行,M-Cdk主导的部分和APC/C主导的部分的分界点为中期和后期的分界点。
- 在G2期,M周期素的转录和合成增多,但此时Cdk被Wee1酶磷酸化抑制,因此M-Cdk会积累,但无活性。(在动物胚胎细胞中则M周期素转录和合成量不变)
- 通过一个尚不清楚的机制,在G2期末,Cdc25酶的活性突然上升,将Cdk酶脱磷酸化激活,引起M-Cdk活性突然上升,细胞进入M期。
- 激活的M-Cdk会抑制Wee1酶和激活Cdc25酶,进一步促进M-Cdk活性上升。
- M期初,还有两种重要的激酶:Polo激酶、Aurora激酶。
- 前期:
- 植物细胞的细胞核要先移动至细胞中央,有时称为预前期(Preprophase)。
- 为防止姐妹染色单体分离时被拉断,细胞投入巨大的能量浓缩染色质,形成染色体。
- 浓缩染色质是通过凝缩蛋白(Condensin)实现的。(图17)
- 染色质浓缩的同时,拓扑异构酶II将两条染色单体整理为易于分开的结构,必要时去除少量黏连蛋白。
- 中心体周围开始组织纺锤丝,植物细胞中染色体周围开始组织纺锤丝。(见下文)
- 去除中心体的细胞仍能正常分裂。
- 中心体中γ-tubulin含量增加,称为中心体成熟(Centrosome Maturation)。
- 纺锤丝由微管组成,MTOC处是负极,分三种:(图18)
- 极间微管(Interpolar Microtubule):两个MTOC伸出的极间微管的正极相互结合。
- 动粒微管(Kinetochore Microtubule):微管的正极和染色体的动粒(Kinetochore)结合。
- 星状微管(Astral Microtubule):微管的正极与细胞膜结合,用于固定。
- 与纺锤丝相关的马达分子有5种:(图19)
- Kinesin-5有2个马达亚基,分别与极间微管的2条微管结合,并分别向正极移动,它把2条微管推开。
- Kinesin-14有1个马达亚基和1个固定的与微管结合的亚基,它也与极间微管的2条微管结合,把2条微管拉紧。
- Kinesin-4和Kinesin-10有1个马达亚基,同时与染色体结合,负责将染色体推至微管末端。
- Dynesin有1个马达亚基,同时与细胞膜下的肌动蛋白结合,负责将MTOC拉至细胞两极。
- Aurora和Polo激酶可磷酸化中心体的蛋白质,促进中心体成熟。
- M-Cdk和Aurora激酶可磷酸化Kinesin-5蛋白,促进中心体分离。
- 核仁消失。
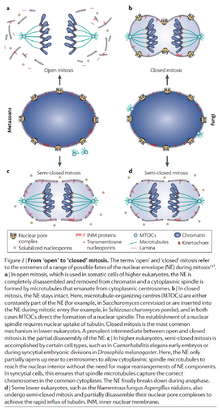
- 前中期:
- 动物和植物细胞中核膜解体或半解体。(被认为是M-Cdk磷酸化核孔复合体引起的)
- 真菌的中心体在核膜上,核膜不解体或半解体。(图16)
- 解体的核膜融入内质网。
- 纺锤丝开始和染色体上的着丝粒(Kinetochore)相连。
- 微管的不稳定性增大,微管骨架重组加速,适应于捕捉姐妹染色单体。(此现象在前期开始,但在前中期和中期较为明显)
- 染色体附着有GEF蛋白(Guanine Nucleotide Exchange Factor),它促使附近的Ran蛋白与GTP结合而不是GDP。
- Ran-GTP能分解某种蛋白复合体,释放微管稳定因子,从而使染色体附近的微管骨架稳定,易于捕捉姐妹染色单体。
- 无中心体的细胞(包括植物细胞)主要依靠染色体稳定微管的能力组织微管,组织好的微管随后被一些马达分子接管。
- 纺锤丝和染色体相连后,染色体所受拉力有三种:
- 动粒处的马达分子的拉力。(这一拉力不消耗ATP,其能量来自微管解体释放的能量,最终来源于微管聚合时水解的GTP)
- 中心体处微管解体并拉动后面的微管,称为微管流动(Microtubule Flux)。
- Kinesin-4和Kinesin-10将染色体向纺锤丝末端推,称为极风(Polar Wind)。
- 中期:
- 当所有的着丝粒都与两极的纺锤丝相连时,细胞进入中期。
- 动物细胞的着丝粒可与10-40根微管相连,而酵母细胞的着丝粒只能与1个相连。
- 纺锤丝与着丝粒外部的杆状蛋白复合体Ndc80相连。
- 染色体一般先与纺锤丝侧面接触,然后被马达分子推至纺锤丝末端。
- 染色体一般不会一次就正确地与两极的纺锤丝相连,而是经多次尝试,不正确的连接方式高度不稳定。(图20)
- 着丝粒内部有Aurora激酶,能磷酸化Ndc80,促使Ndc80和微管分离。
- 若着丝粒与纺锤丝正确相连,纺锤丝的拉力使Ndc80远离Aurora激酶,防止它被磷酸化,所以正确的连接方式稳定。(图21)
- 最终,染色体因受到两极大小相近的拉力而静止在细胞中央,所有染色体组成的平面称为赤道板(Metaphase Plate)。
- 后期:
- 未正确与纺锤丝相连的着丝粒会引起Mad2蛋白构象改变。
- 构象不正确的Mad2蛋白会抑制Cdc20蛋白,从而抑制APC/C复合体。
- 当所有染色体正确排列在赤道板上时,Cdc20抑制解除,APC/C被激活。
- APC/C降解保全素(Securin),激活之前与之结合的分离酶(Separase)。
- 分离酶降解黏连蛋白,从而分离姐妹染色单体。(图22)
- 一般以所有姐妹染色单体分离为后期开始。
- APC/C降解S和M周期素。
- 秋水仙碱(Colchicine)、长春碱(Vinblastine)能阻碍微管聚合,阻止细胞进入后期,也可阻止姐妹染色单体分离,使染色体加倍。
- 后期可分为A和B两部分,略有重叠。
- 后期A:微管正极解散,将染色体向两极拉动。
- 后期B:中心体自身向两极移动,将染色体向两极拉动。
- 末期:
- 内质网上分离出核膜,将两极的染色体包裹成核。
- 核仁重新出现。
- 胞质分裂:
- 在后期,细胞中间开始出现收缩环(Contractile Ring)。
- S和M周期素降解能促进收缩环形成,保证它在正确时间形成。
- 收缩环主要由肌动蛋白、肌球蛋白II组成,还有其它结构和调控蛋白。
- 收缩环与细胞膜上的蛋白结合,逐渐收缩。
- 一般以收缩环收缩至细胞膜突然出现明显凹痕(胚胎学中称为卵裂沟(Cleavage Furrow))作为胞质分裂开始,一般在末期。
- 收缩环收缩的同时,蛋白不断减少,保持粗细不变。
- 细胞膜面积增大的部分由来自高尔基体的囊泡提供。
- 最后,收缩环解散,两个子细胞中间通过中体(Midbody)相连。(图24)
- 一部分微管不随中心体移动至子细胞中,留在中体形成中心纺锤体(Central Spindle)。
- 中体仍会继续收缩,至两个细胞完全分离,中心纺锤体一般移入其中一个子细胞。
- 收缩环由RhoA激酶启动,见图25。
- 收缩环的位置确定有三种理论,见图26:(多数细胞使用的方式是这三种理论的混合体)
- 星状微管刺激(Astral Stimulation):在细胞膜上的固定点位于细胞中间的星状微管,能与促进收缩环形成的调控蛋白结合。
- 中心纺锤体刺激(Central Spindle Stimulation):极间微管与若干信号分子结合,其中一种能激活RhoA。
- 星状微管松弛(Astral Relaxation):星状微管对细胞膜有拉力,而细胞中间处受到的拉力最小,特定蛋白能感受此拉力。
- 酵母细胞在G1期末有胞裂蛋白(Septin)聚集在细胞特定部位,提前确定了收缩环位置。
- 植物细胞不通过收缩环分裂细胞,而是在细胞中间构建新细胞壁。
- 植物细胞在后期,部分微管在细胞中间构建成膜体(Phragmoplast)。
- 来自高尔基体的囊泡沿着成膜体的微管在细胞中间形成细胞板(Cell Plate)。
- 囊泡的主要成分为果胶(Pectin)、少量糖蛋白,无纤维素。
- 细胞板最终成为中胶层(Middle Lamella)。