{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
总论[ | ]
- 对植物发育有影响的环境因素有:温度、光照、水分、营养、重力、与其它生物的交互,其中最主要的是温度和光照。
温度的影响[ | ]

{kind=link}
图1:水毛茛的叶 J. E.(Jed) and Bonnie McClellan © California Academy of Sciences
- 在恒温条件下,植物有可生长的温度范围,将生长速度对温度作图,有一个最适温度的峰,通常在30℃附近。
- 植物最适应的环境,其温度是周期性变化的,此现象称为温周期(Thermoperiodism)。
- 温度会影响器官发育的结果,以水毛茛(Water Buttercup,学名Ranunculus aquatilis)为典型:
- 水毛茛是水生植物,它有两种叶子:浸入水下的呈羽状全裂(Filiform Disected),而露在水上的无缺刻。
- 只有一天中的最低水温在8~18℃时,才能发育出羽状全裂的叶;如果升高水温,则叶呈无缺刻形态。
- 如果对水毛茛施用脱落酸,则水下的叶也会发育出无缺刻形态;如果光照强烈,蒸腾作用旺盛,脱落酸生成也会增加,形成水下无缺刻的叶。
- 牵牛花的花色部分由花芽发育时的温度决定,低温会促进色素合成。
- 热带雨林中,咖啡、兰花、部分竹子物种会在下雨后集体开会,其决定因素是突然而短暂的低温。
- 适当低温能打破种子休眠,促进种子萌发,称为分层现象(Stratification):
- 对大多数植物,以0~5℃为宜,有些高山植物要求0℃以下,被称为Frost Germinator。
- 有些物种必须在低温下才萌发,如白蜡;其它则低温只有加速作用。
- 有些物种只有完整的种子需要低温才萌发,则将胚单独培养则不经低温也萌发,如西克莫槭(Acer pseudoplatanus)
- 棉花、大豆、高粱反而需要高温才能萌发。
- 草地早熟禾(Poa pratensis)在15~18℃之间波动的环境中萌发最适。
- 低温对干种子无效。
- 山楂(Crataegus)、栒子(Cotoneaster)的种子落地后,只能在第二个春天萌发,因为它们的种皮很厚,第一个春天时吸水不足。
- 铃兰(Convallaria)、玉竹(Polygonatum,又称黄精)、延龄草(Trillium)的胚根在第一个春天生长,上胚轴则要到第二个春天生长。
- 杏、牡丹不需要低温就能萌发,但上胚轴只有在经历低温后才生长。
- 低温对种子休眠的影响很复杂,可能是调节了某些激素的产量或酶的活性。
- 对种子施用赤霉素(Gibberellin)通常能免去对低温的要求,但低温对赤霉素生产的影响未知。
- 低温能促进某些物种开花,称为春化作用(Vernalization):
- 只要一片叶子接受低温,整株植物开花都会被促进,因此认为低温引起植物产生了一种激素,被称为春化素(Vernalin)。
- 春化素的化学本质未知,有人根据其对植物的影响,认为它就是另一种猜测存在的激素促花素(Florigen,见下文),促花素的化学本质也是未知的;也有人认为它是赤霉素,因为用赤霉素处理植株能跳过春化作用开花,但是赤霉素已被证明不是促花素。
- 需要春化作用的物种:
- 冬性一年生植物:冬小麦、冬黑麦、冬大麦(统称为冬性谷物)、春葶苈(Erophila verna)、婆婆纳(Veronica agrestis)、勿忘草(Myosotis discolor)、白芥(Sinapis alba)。
- 二年生植物:甜菜(Beta vulgaris)、芹菜(Apium graveolens)、包菜、萝卜、白菜(和其它十字花科植物)、胡萝卜、葱、蒜、荠菜、百合、鸢尾、郁金香、风信子、二年生天仙子(Hyoscyamus niger)、洋地黄(Digitalis purpurea)。
- 二年生天仙子需要在春化作用过后又经历一天长日(日照时间大于一定时长)才能开花。
- 多年生植物:樱草、紫罗兰、桂竹香、菊花、紫菀、康乃馨、英国黑麦。(它们必须每年接受一次春化作用)
- 所有以上物种都可以在枝繁叶茂时接受春化作用;有些物种在种子阶段就能接受春化作用,如甜菜、白芥、冬性谷物,它们的特点:
- 低温只对开花起促进作用,没有低温也能开花。
- 春化作用是逐步进行的,在完成之前,加高温(40℃)可逆转春化作用,称为脱春化作用(Devernalization),但完成后就不可逆转。
- 分层现象和春化作用的意义:防止植物过早发育,然后在冬天被冻害。
光照的影响[ | ]
{kind=link}
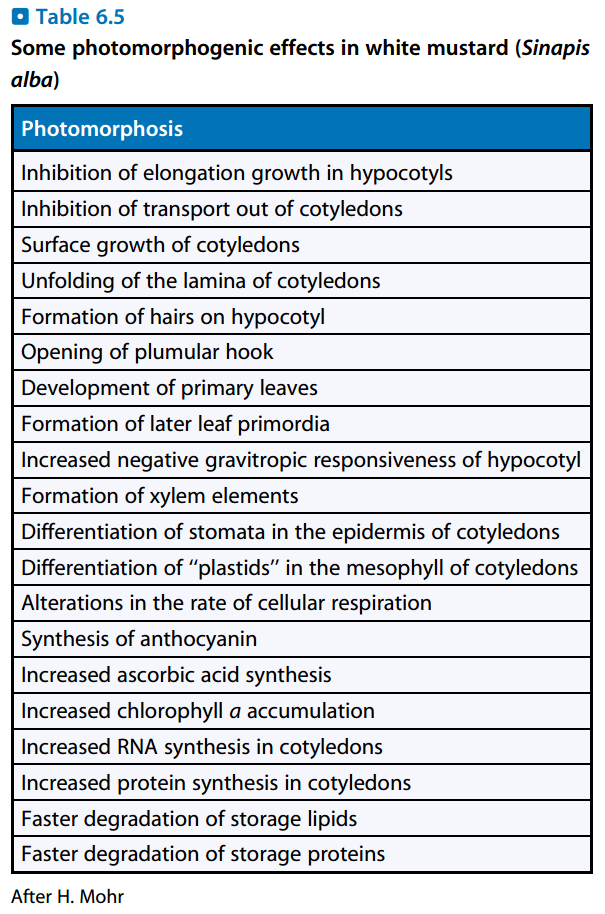
图2:白芥中的光形态建成

{kind=link}
图3:长日植物、日中植物、短日植物

{kind=link}
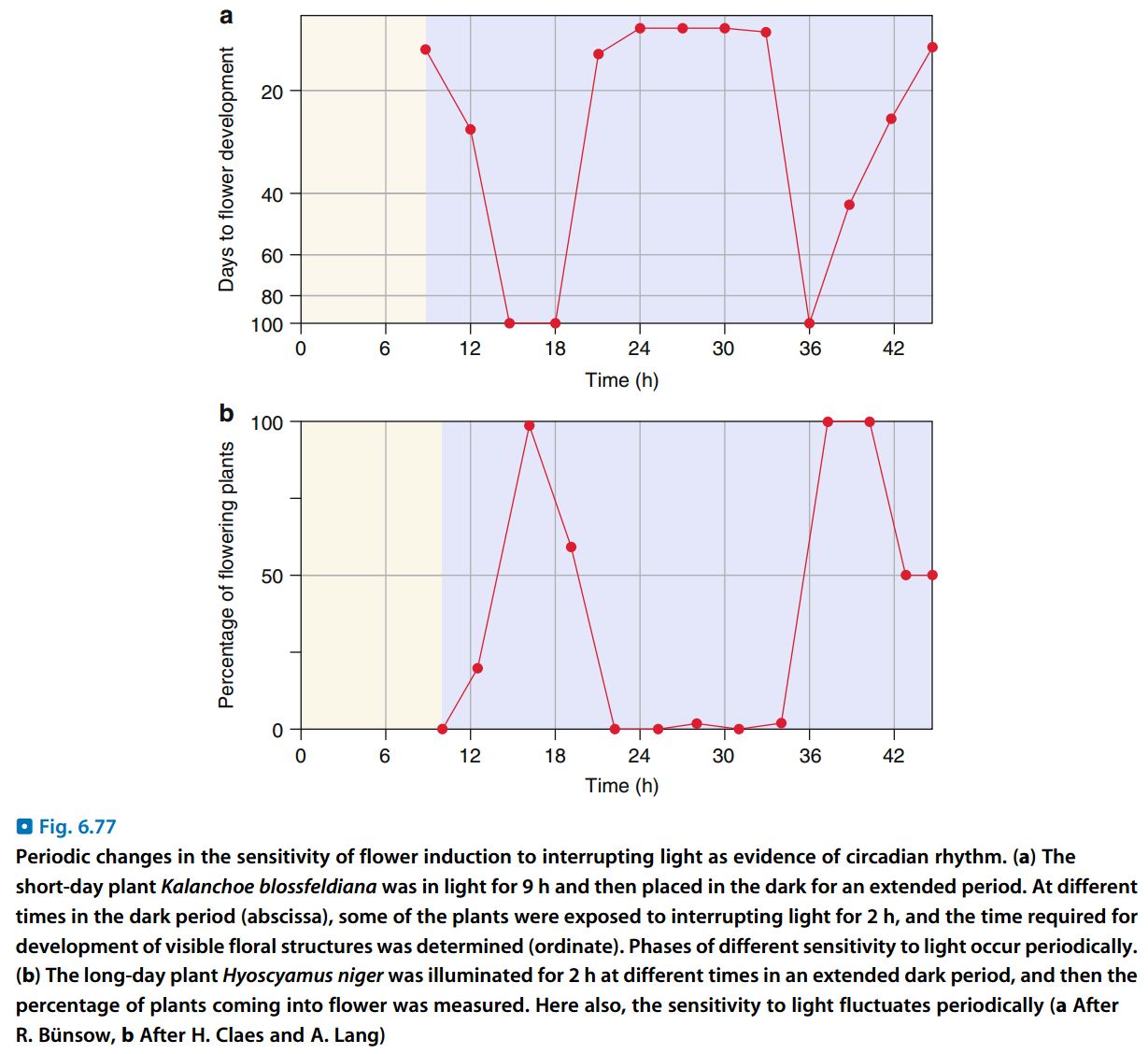
图4:植物在长夜中不同时段对打断的光照敏感度不同

{kind=link}
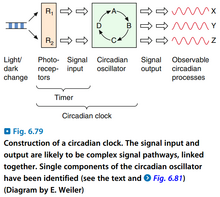
图5:生理时钟

{kind=link}
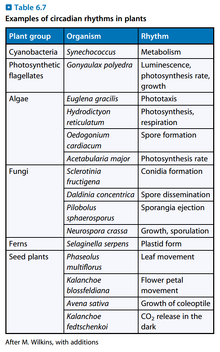
图6:各类植物中的生理时钟现象


{kind=link}
图6:植物的各类光感受器(Cytochrome即文中的Cryptochrome)


{kind=link}
图7:光敏素

{kind=link}
图8:光敏素引起的各类生理反应
{kind=link}
图9:植物光感受器的进化
- 对于大多数低等植物,在光下和黑暗中发育的结果类似,但在被子植物中,光对植物发育有显著影响,称为光形态建成(Photomorphogenesis)。
- 将光下生长的植株移至暗处,它会变黄并过度生长,称为黄化作用(Etiolation);将种子直接在暗处培养也有类似结果,称为暗形态建成(Skotomorphogenesis)。
- 将黄化的植株重新移至亮出,又会脱黄化(De-etiolation)。
- 喜阳植物移至阴处,也会有部分黄化植株的特征。
- 黄化植株的特征:
- 节间长度大幅增长,有时还包括叶柄、叶片。
- 形成胚芽钩(Plumular Hook),萌发的芽顶部是弯曲的。
- 不形成维管束,细胞中各类色素的合成也被抑制。
- 茎和叶非常柔软。
- 如果使拟南芥的COP基因或DET基因失活,则它在暗中也进行光形态建成。
- 暗形态建成的意义:使植物集中营养生长到有光的地方。
- 光照影响细胞或组织的极性,也就是背腹侧形态不同,它实际上是由光照强度的差别造成的。
- 接受阳光较多的叶,如北半球的树的向南的叶,或树靠外侧的叶,其栅栏组织更厚,Rubisco酶更多,可溶蛋白含量更高;一些物种中光照还会影响器官的形态。
- 低等植物中的光形态建成现象:
- 衣藻(Chlamydomonas)只有在光下才生成生殖细胞。
- 蕨类植物在暗中或红光下只形成原丝体,必须加入蓝光才能继续发育。
- 真菌的发育也需要蓝光,个别物种还需要紫外线。
- 苔通过光照决定原叶体哪一面是“背面”,在光照强的一面长出性器官,一些蕨类植物也有类似现象。
- 植物在一天之内受到的光照时长称为光照期(Photoperiod),它也会影响植物发育:
- 一般来说,只要光照强度达到10-3 W m-2,就算作光照期,具体的光照强度几乎没有影响,因此特定情况下满月也能算入“光照期”。
- 光照期长度对发育活动的影响,可基本分类为:长日现象(光照期必须大于一定时间才发生)、短日现象(光照期必须小于一定时间才发生)。(光照没有影响,称为日中现象)
- 这里所说的“一定时间”,称为临界日长(Critical Day Length)。
- 有些发育活动的决定因素是光照期长度的变化,称为定量长日现象、定量短日现象。(对应地,上述两种基本分类现在称为定性长日现象、定性短日现象)
- 有些发育活动需要先接受一定时间的长光照期,然后接受一定时间的短光照期,称为长-短日现象;有些又恰好反过来,称为短-长日现象,它们被统称为中日现象(注意与日中现象区分)。
- 光照期影响最显著的活动是开花,因此按光照期对开花的影响将植物分为“长日植物”“短日植物”“日中植物”。
- 一个光照期加上一段时间的黑暗,称为一个光周期,发育活动不一定经历一个满足要求的光周期就能发生,有些需达25天才能发生,如长叶车前草(Plantago lanceolata,长日植物)。
- 对于短日植物,光照时间太短也不行,因为不能进行光合作用,营养不够。
- 植物的光照类型与其地理分布有关:
- 热带几乎没有长日植物或定量植物,因为那里的日照长度几乎没有变化,总是约12 h。
- 高纬度植物几乎都是长日植物,因为它们必须在夏天开花,秋天结果,否则就会被冻害。
- 中纬度则各类植物都有,其分布与旱雨季有关。
- 印度、中国、中美的旱季在冬天,那里多为短日植物。
- 地中海、中东、中亚的旱季在夏天,那里多为长日植物。
- 实验表明,只要植株的一片叶子,甚至叶子的一部分,接受了满足条件的光照期,就能进行发育活动,如将长日植物置于恒定光下,将一片叶子遮光,就能引起开花。
- 因此猜测光周期能引起植物产生一种激素,被称为促花素(Florigen),其化学本质仍是未知的。
- 将长日植物和短日植物都放在长日的光周期下,将开花的长日植物嫁接到短日植物上,短日植物也开花,因此促花素应当是在各植物中一致的物质。
- 促花素在顶端分生组织起作用,它的运输速度很慢,因此认为它是在共质体中运输。
- 若对于短日植物,置于黑夜足够长的光周期中,并用光照短暂(1分钟)打断黑夜,则也不会开花;用短暂黑暗打断光照则不会有效果,因此有人主张“长日植物”应称为“短夜植物”,“短日植物”应称为“长夜植物”。(但是对于长日植物置于长夜的光周期中,打断黑夜的光照必须有几个小时)
- 在黑夜中的不同时段,植株对打断的光照的敏感度不同,见图4。
- 现在的分子生物学认为,光照对植物的影响是通过调控植物自身的生理时钟(Physiologic Clock)起作用的。
- 生理时钟的特点:
- 如果保持所有外界条件恒定,则它仍能稳定地运行1~2周,在低等植物中长一些,可达几个月;随着时间推移,各种分子层面的周期性变化的幅度会减弱。
- 虽然恒定条件下它能稳定运行,但它的周期通常不是24 h,而是稳定在一个略长(可达30 h)的长度,有时也会略短。
- 温度对生理时钟的影响极小,因此认为存在一种还未发现的机制,能补偿因温度升高加快酶促反应造成的生理周期缩短。
- 生理时钟被认为在进化中反复出现了多次,对它的具体的分子机制的研究的模式物种有:果蝇(Drosophila)、粗糙链孢菌(Neurospora crassa)、聚球藻(Synechococcus,一种蓝藻)、拟南芥(Arabidopsis thaliana)。
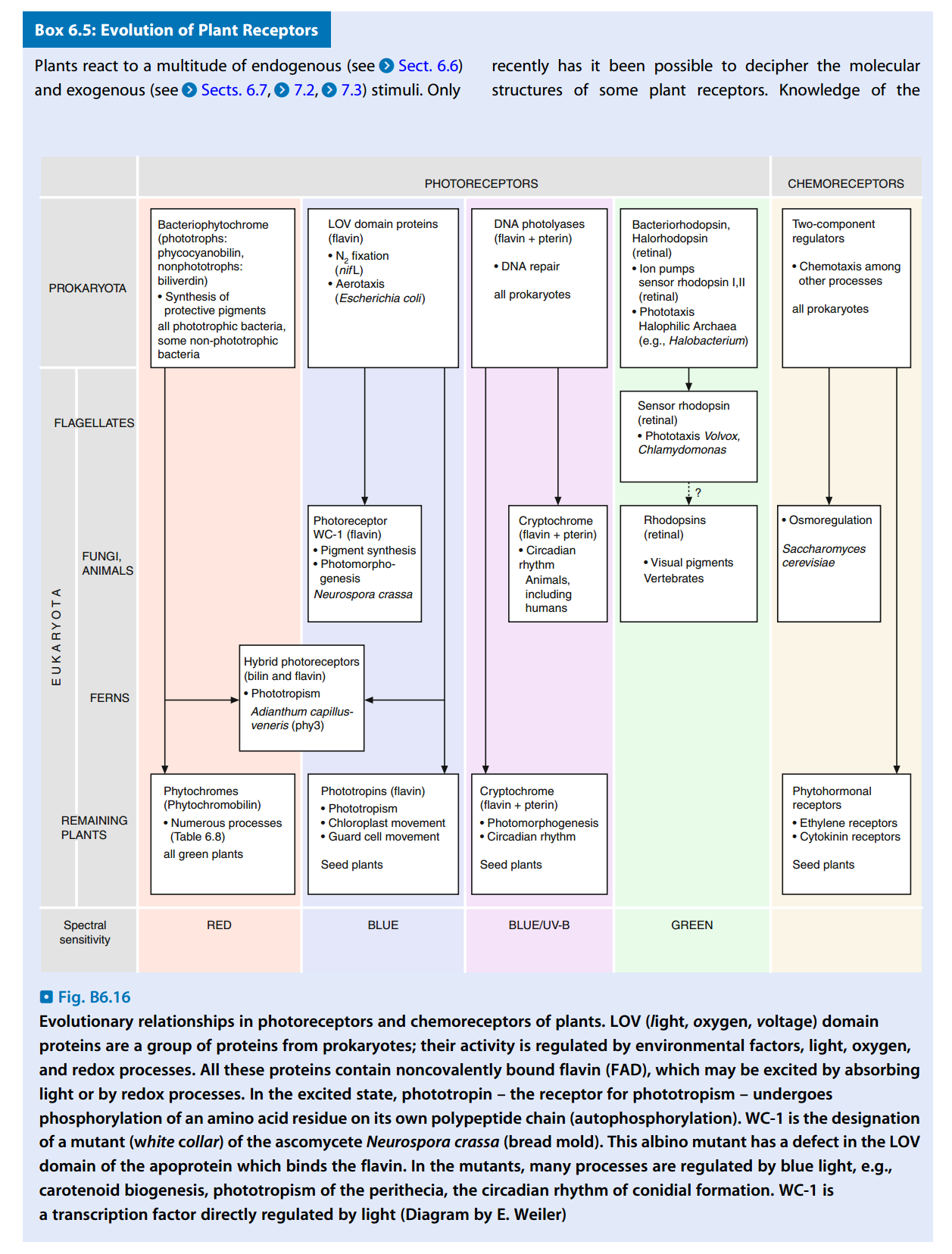
- 植物的光感受器:
- 真菌只对520 nm以下的光敏感(蓝光和紫外线)。
- 绿色植物不只对蓝光敏感,红光对它的作用更大。
- 由光感受器的信号传导引起的不可逆变化,称为光分化(Photodiffertiation);引起的可逆变化,称为光调节(Photomodulation)。
- 目前发现的植物光感受器有:光敏素(Phytochrome,分I类和II类)、隐色素(Cryptochrome)等,见图6。
- 光敏素的发色团是植物色素(Phytochromobilin),结构上属于四吡咯(Tetrapyrrole),由叶绿体合成。
- 植物色素和光敏素多肽在细胞质中结合,通过二硫键共价连接,此连接是光敏素多肽催化的。
- 在黑暗中产生的光敏素,其吸收峰在667 nm处,但实验中常用660 nm的红光激活它,因此这种光敏素被记为P660或PR。
- PR被刺激后,植物色素的一个双键发生异构反应,光敏素的吸收峰移动至730 nm的远红光处,因此它被记为P730或PFR。
- PFR是有信号传导活性的光敏素构象,用远红光照射PFR,它又能恢复到PR的状态。
- PR和PFR的光吸收峰有重叠,用660 nm的红光长时间照射至平衡时,有20%的光敏素仍是PR;用730 nm的远红光照射至平衡时,有2.5%的光敏素仍是PFR。
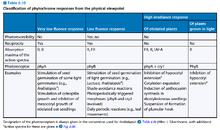
- 引起不同的生理反应所需的PFR的量是不同的,据此将生理反应分为VLFR(Very Low Florescence Response)、LFR(Low Florescence Response)、HIR(High Irradiance Response)。
- VLFR反应一旦启动,用红光照射是不可逆的。
- VLFR反应和LFR反应一般适用乘积规则,即反应强度与光照时间和光照强度的乘积成正比,也就是可以用较弱但较长的光替代较强但较短的光。
- HIR反应的强度主要和光照强度成正比,与光照时间无关。
- 被子植物既有I类光敏素又有II类光敏素,所有其它植物只有II类光敏素。
- I类光敏素在光下会迅速被泛素系统降解,同时其合成被抑制;II类光敏素在光下和黑暗中都稳定。
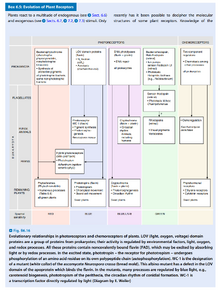
- 在细菌中也发现了类似光敏素的蛋白,甚至在不进行光合作用的细菌中也有。
- 光合细菌中光敏素通过半胱氨酸与一种类似植物色素的物质连接;在非光合细菌中它通过组氨酸与胆绿素(Biliverdin,一种血红素降解产物)连接。
- 隐色素的发色团是蝶呤(Pterin)和FADH,它在结构上类似于用于修复DNA的光裂解酶(Photolyase),它的吸收峰在蓝光和紫外线。
- 隐色素通常起辅助光敏素的作用。
- 植物光感受器启动的信号通路目前大部分仍是未知,可能是移动至细胞核中,调节各种转录因子。
植物的向性运动和感性运动[ | ]

{kind=link}
图10:植物向光生长的反应光谱


{kind=link}
图11:两次向光反应


{kind=link}
图12:两次向光反应有区别


{kind=link}
图13:有多个光源时,向光弯曲方向遵从平行四边形法则


{kind=link}
图14:离心力对向地性生长的影响

{kind=link}
图15:高等植物向地性生长的生长素模型(根)
- 植物的“运动”通常指器官向特定的方向发育。
- 刺激的方向控制运动的结果,称为向性运动(Tropism);刺激只启动运动,运动的结果只由植物的结构控制,称为感性运动(Nastic Movement)。
- 向性运动又分为:正向性运动(植物朝着刺激来源发育)、负向性运动(植物背离刺激来源发育)、斜向性运动(Plagiotropism,植物发育方向与刺激方向成一角度)、横向性运动(Diatropism,斜向性运动的特例,即发育方向与刺激方向垂直)。
向光性生长[ | ]
- 很多植物的茎和叶柄都有正向光性生长(称为Phototropism)。
- 负向光性生长:出现于气生根、固着根、苔的假根、蕨类的原叶体、葡萄藤、白芥(Sinapis)的胚根。
- 横向光性生长:出现于一些植物的侧枝、叶、藓的原叶体。
- 有些植物的花茎在受精前向光生长,在受精后背光生长。
- 还有一种特殊的向光性生长,称为向阴生长(Scototropism),即总是向着地平线上最暗的方向匍匐生长,出现于热带藤本植物,其分子机制不明。
- 引起向光性生长的光感受器:
- 控制苔藓、蕨类植物的向光性生长的光感受器,被证明就是光敏素,上文已介绍,下文适用于种子植物。
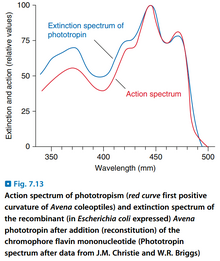
- 根据单色光引起的向光生长速度的差异,发现感受器的吸收峰在蓝光和紫外线区域。
- 目前发现了两种控制向光生长的光感受器:NPH1和NPL1(现更名为PHOT1和PHOT2),它们都有蛋白激酶活性,它们的发色团都是非共价结合的FMN。(它们统称为向光素,Phototropin)
- 向光生长符合LFR反应的特征,即强度与光照强度和时间的乘积相关。
- 在相同时间内提供不同强度的光照,发现随着光强度增加,向光生长反应强度先升高,后降低(称为第一次向光反应),然后又升高,再降低(称为第二次向光反应,此时光照强度已超过自然阳光),接着还有第三次向光反应(没有实际意义)。
- 实际上,直接暴露在阳光下的植株主要是第二次向光反应控制,而埋在土壤中的种子主要受第一次向光反应控制。
- 向光素的下游信号通路不明,但最终是控制生长素(Auxin)的分布,使不受光一侧的生长素较多,细胞分裂较快,从而器官向受光一侧弯曲。
- 胚芽鞘的向光生长反应:
- 对光最敏感的点是尖端,只刺激尖端也会引起向光生长,但刺激基部则不敏感。
- 第一次向光反应时,曲率最大部位先在尖端,后移至基部。
- 第二次向光反应时,曲率最大部位直接在基部。
- 两次向光反应都被证明只有PHOT1参与。
- 有多个光源存在时,胚芽鞘弯曲方向遵从平行四边形法则,见图13。
向地性生长[ | ]
- 正向地性生长:出现于藻类、苔的假根、蕨类的原叶体、种子植物的根。
- 负向地性生长:出现于种子植物的茎。
- 横向地性生长:出现于一级侧根、侧枝、叶、根茎。(二级及以上侧根一般无向地性反应;这些器官有时是斜向地性生长;柳树的侧枝也没有横向地性生长)
- 罂粟花在花苞状态是正向地性生长,将要开花前则变为负向地性生长。
- 很多植物的花茎是负向地性生长,果茎则变为正向地性生长,欧洲百合(Lilium martagon)则恰好相反。
- 感受重力的部位:胚芽鞘中是尖端的叶肉组织;根尖是根冠细胞;茎中是所有还在生长的节间的伸长区。
- 离心力能影响植物的重力感应,见图14。
- 能感受重力的细胞的特点:
- 细胞结构分布很不对称,低密度的(如液泡)都在上部,而高密度的(如细胞核、淀粉体、一些盐的晶体)都在下部。(鱼藻含硫酸钡晶体,被称为平衡石,Statolith)
- 在黑暗中生长而无淀粉积累,或淀粉合成有障碍的植株,其重力感受很弱,但不是完全消失,其高密度颗粒主要是白色体。
- 重力感受细胞似乎是通过抑制一侧细胞分裂起作用的。
- 植物细胞感受重力的方式尚不确定,有三种学说:
- Topographic学说:细胞直接感受细胞结构的分布。
- Kinetic学说:细胞感受各细胞结构的运动情况。
- Deformation学说:细胞的各结构在运动时挤压特定关键结构,被细胞感受。
- 可能挤压内质网或高尔基体,控制其释放囊泡的方向。
- 可能挤压细胞骨架,再传感到其它细胞器。
- 可能直接挤压细胞膜,这是目前接受最多的模型。
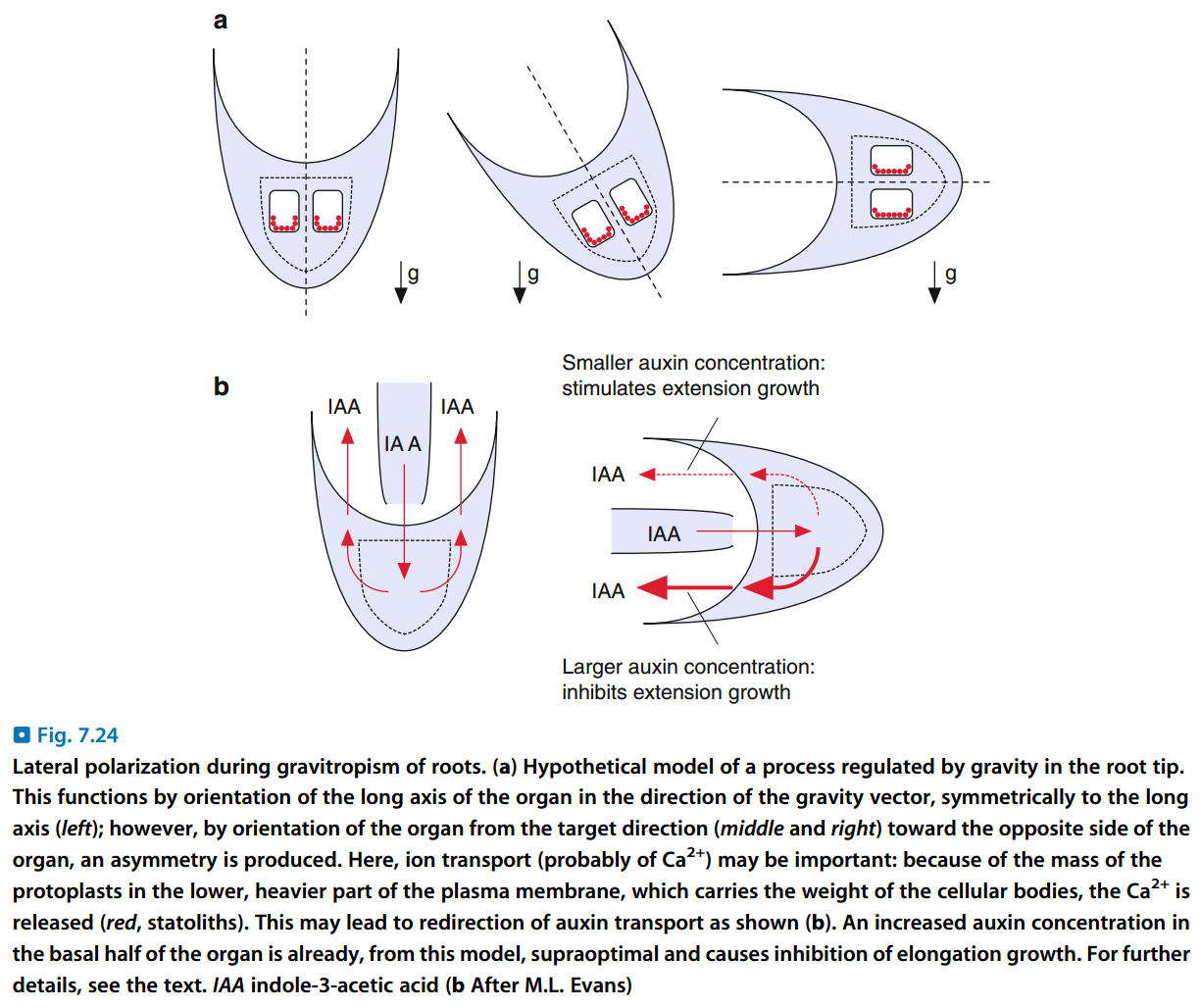
- 根中的重力感受要求细胞外有一定浓度的Ca2+。
- 高等植物的向地性生长适用生长素模型:
- 无论根中还是茎中,生长素都集中到重力所指的一侧,即下侧。
- 茎中,下侧生长被促进,上侧生长被抑制,结果向上弯曲,呈负向地性生长。
- 根中,高浓度的生长素抑制了下侧细胞分裂,而上侧正常分裂,结果向下弯曲,呈正向地性生长。
- 侧枝的横向地性生长是负向地性生长和偏上性生长(Epinasty)共同作用的结果。(偏上性生长是侧枝上侧的细胞分裂加快的现象,是植物自发的生理过程,与外界刺激无关)
其它向性运动[ | ]
- 其它向性运动有:向触性生长(Thigmotropism)、向化性生长(Chemotropism)。
感性运动[ | ]
待编写