{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
总论
- 病毒没有细胞结构,不能独立生存和复制,必须借助宿主细胞的DNA和蛋白质合成器。
- 病毒的基本结构包括蛋白质外壳和核酸,一半的病毒的外壳外有脂双层膜。
- 蛋白质外壳内可以有一些酶。
- 病毒的分类工作由国际病毒分类学委员会(International Committee on Taxonomy of Viruses,ICTV)进行。
- 病毒的分类级别只有目、科、亚科、属、种。
- 大部分病毒没有被归入某个目中。
- 有些病毒的基因组不含全部自身需要的蛋白质,需要从其它病毒借用,称为缺陷病毒(Defective Virus)。
- 原核生物和真核生物差异巨大,所以存在细菌和古菌都能感染的病毒,但不存在原核生物和真核生物都能感染的病毒。
病毒的结构
{kind=link}
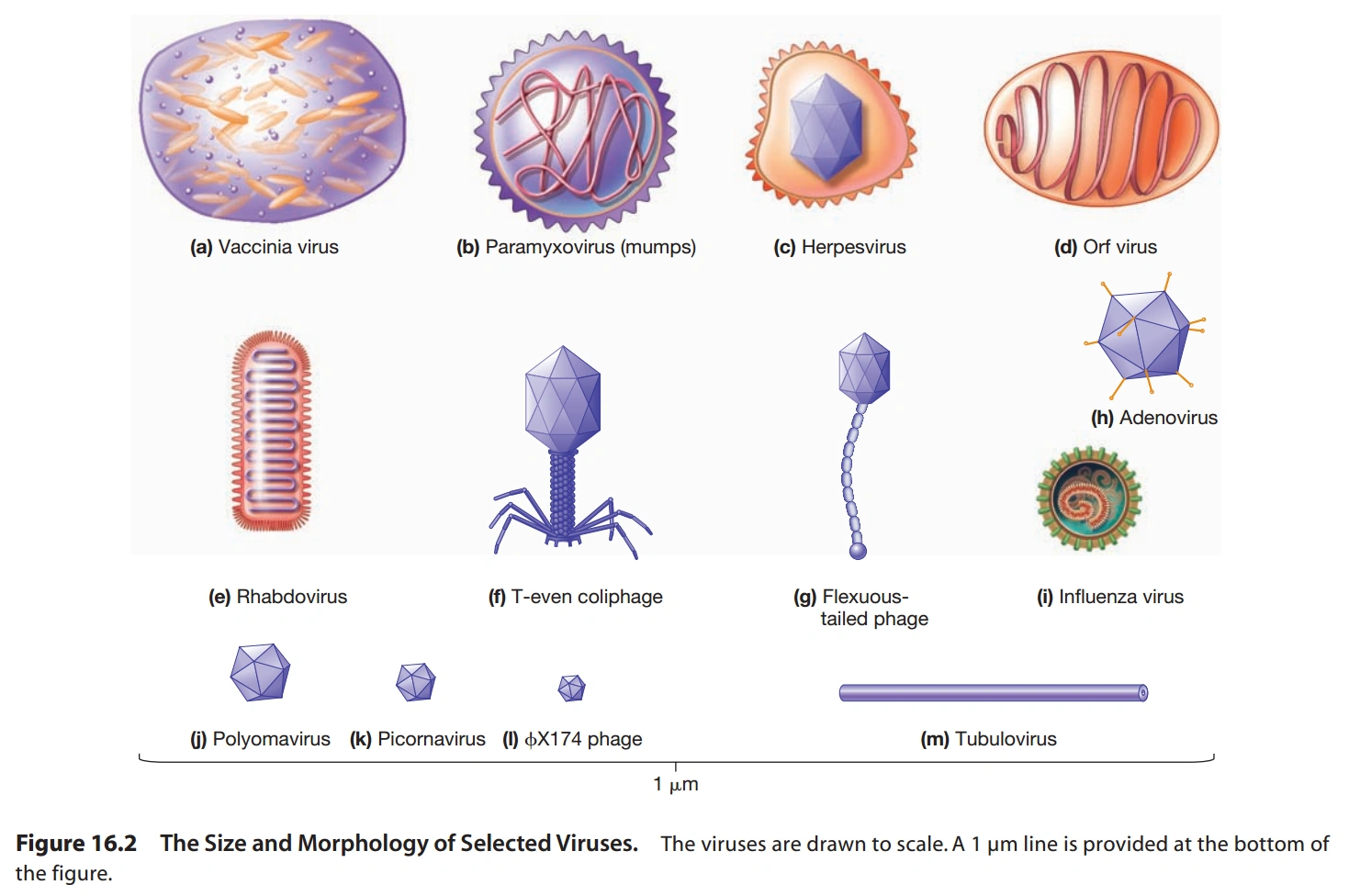
图1a:病毒的形状和大小有很多种
{kind=link}
图1b:病毒的核酸的形态有很多种
{kind=link}
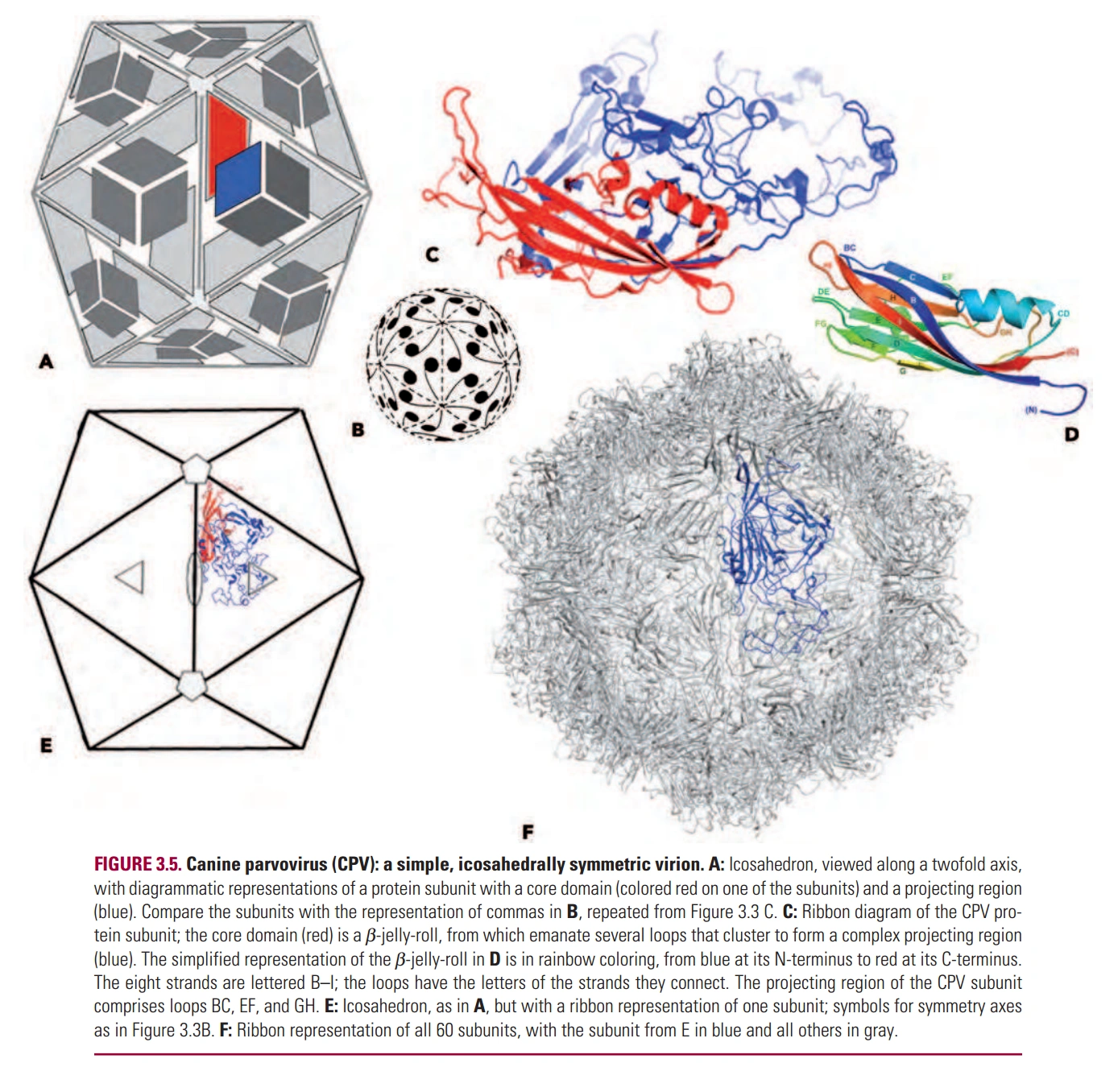
图2:细小病毒的蛋白质外壳

{kind=link}
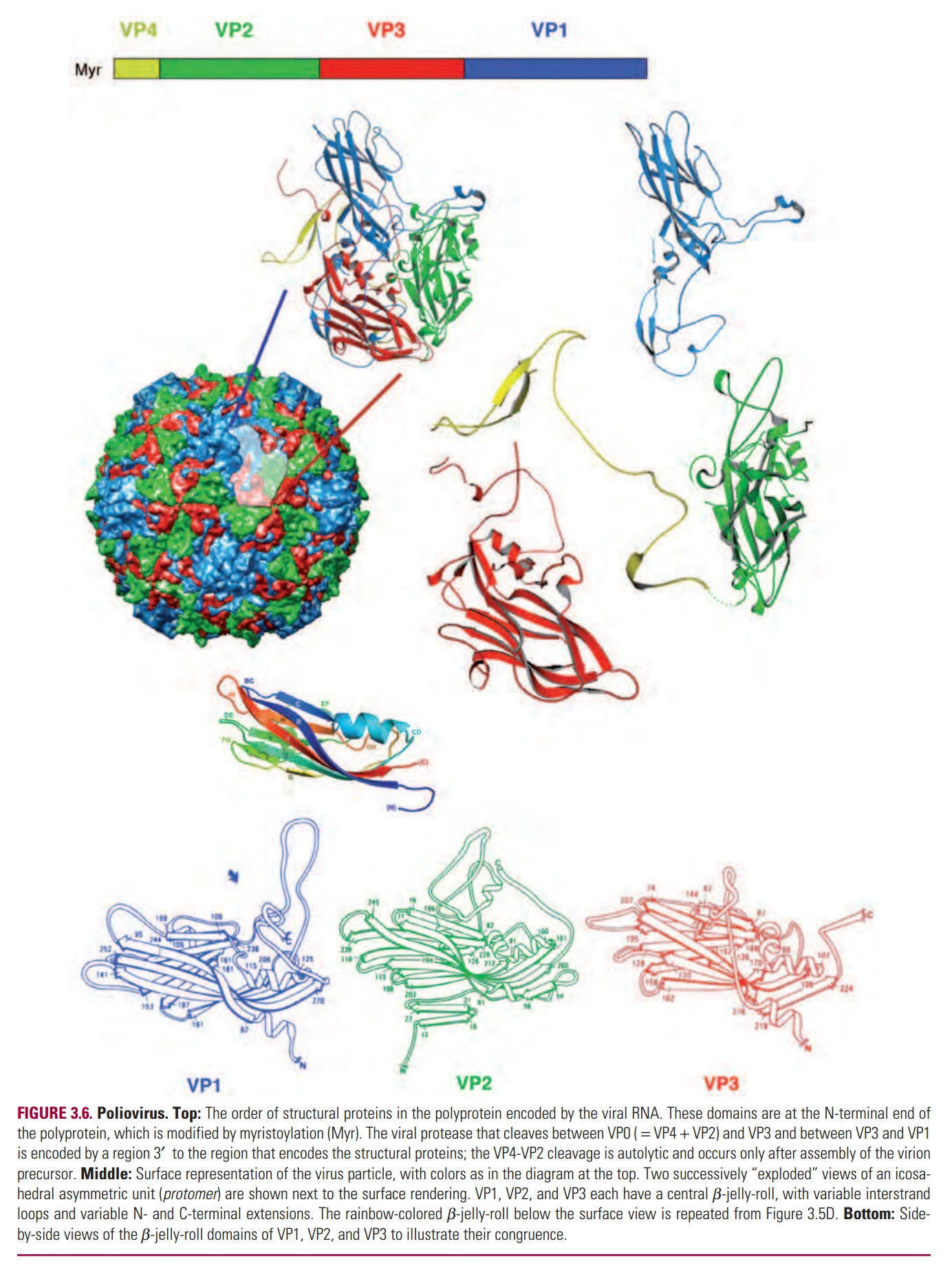
图3:小核糖核酸病毒的蛋白质外壳

{kind=link}
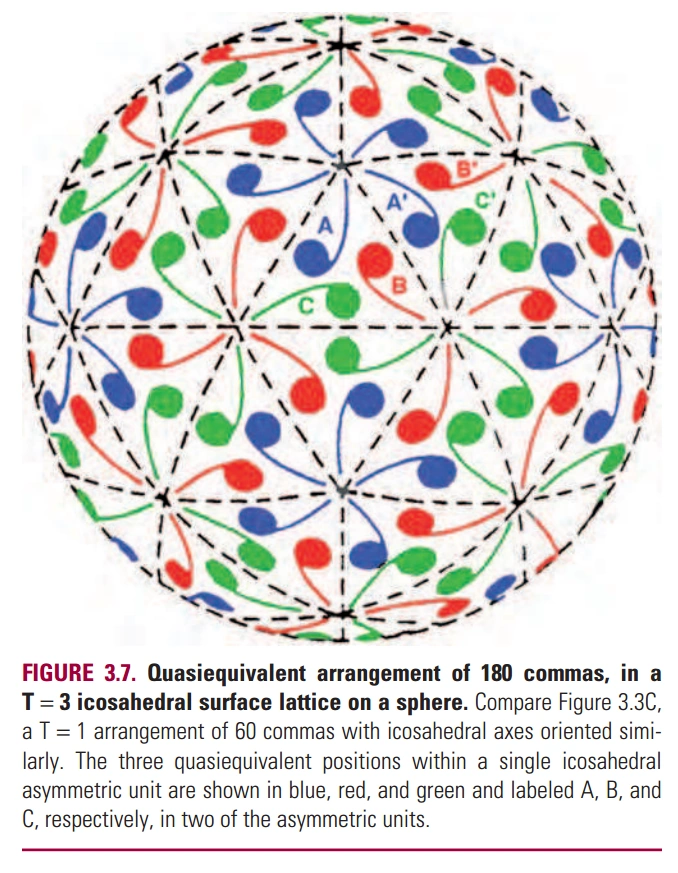
图4:Quasi-equivalence外壳结构
- 传统的观察病毒的方法是用电子显微镜观察被感染的细胞。
- 较大的病毒颗粒可用电子显微镜直接观察。
- 上述方法的缺陷是清晰度较差。
- 用乙酸铀酰(又称乙酸双氧铀,Uranyl Acetate)、磷钨酸钾(Potassium Phosphotunstate)等物质负染病毒颗粒后观察,清晰度较好,但不易辨认内部结构。
- 将病毒颗粒快速降温至液氮的温度,用电子显微镜观察可看到较清晰内部结构,此技术称为冷冻电子显微镜(Electron Cryomicroscopy)。
蛋白质外壳
- 病毒的基因组很小,能编码的蛋白质也很少,所以蛋白质外壳总是由大量的少数几种亚基组成。
- 因为蛋白质外壳亚基种类少,所以外壳结构总是高度对称。
- 虽然结构高度对称,但形状和大小仍有很多种,如图1-4。(大多数病毒是近似正二十面体)
- 基因组最小的病毒,外壳只有一种亚基,如细小病毒科(Parvoviruses)。
- 小核糖核酸病毒(Picornavirus)的外壳由3种亚基组成,每种60个。
- 番茄丛矮病毒(TBSV)、诺瓦克病毒(Norwalk Virus)等的外壳结构较复杂,如图4。
核酸
- 病毒的核酸非常多样。
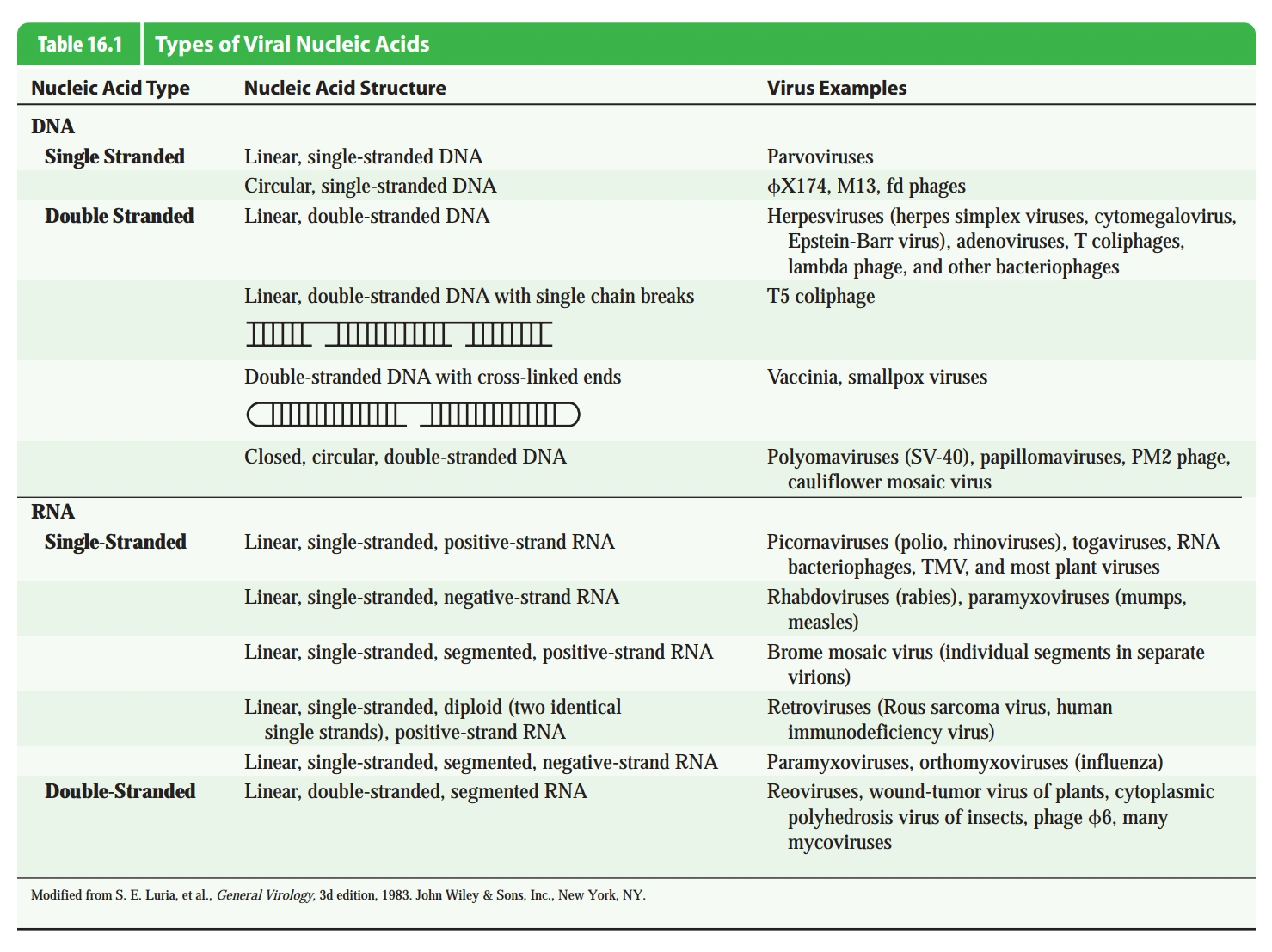
- 按核酸类型分,病毒的核酸为DNA或RNA。
- 按链数分,病毒的核酸既可以为单链,也可为双链;还可为既有单链部分,也有双链部分。
- 按形状分,病毒的核酸既可是线形也可是环形。
- 单链(非反转录,见下文)病毒,根据其核酸的序列和用于蛋白质合成的mRNA的序列是否相同,分为正链(相同)病毒和负链(不相同)病毒。(一般只对RNA病毒作此分类,但对DNA病毒此概念也适用)
- 病毒蛋白质外壳内可以有不止一条核酸。
- 双链DNA和单链RNA病毒较常见。
- 有些病毒在复制过程中从RNA产生DNA,称为反转录病毒(Retrovirus)。
- 反转录病毒只有双链DNA和单链RNA(不分正负链)两种。
- DNA病毒的基因组可以含脱氧尿嘧啶。
- 大多数噬菌体是DNA病毒(双链居多),也有双链和单链正链RNA病毒。
- 所有古菌噬菌体都是DNA病毒,几乎都是线性双链DNA病毒。
脂双层膜
- 脂双层膜在动物病毒中最常见,植物病毒次之,细菌病毒中不常见。
- 脂双层膜上的脂质和糖类一般直接来自宿主细胞,蛋白质由病毒的基因组编码。
- 脂双层膜上的蛋白质在电镜照片下明显突出,称为突刺(Spike)或包膜粒(Peplomer)。
- 一般,脂双层膜的形状可变,称为多形性(Pleomorphism)。
- 狂犬病毒的脂双层膜与蛋白质外壳紧密相连,不具多形性。
病毒的分类

{kind=link}
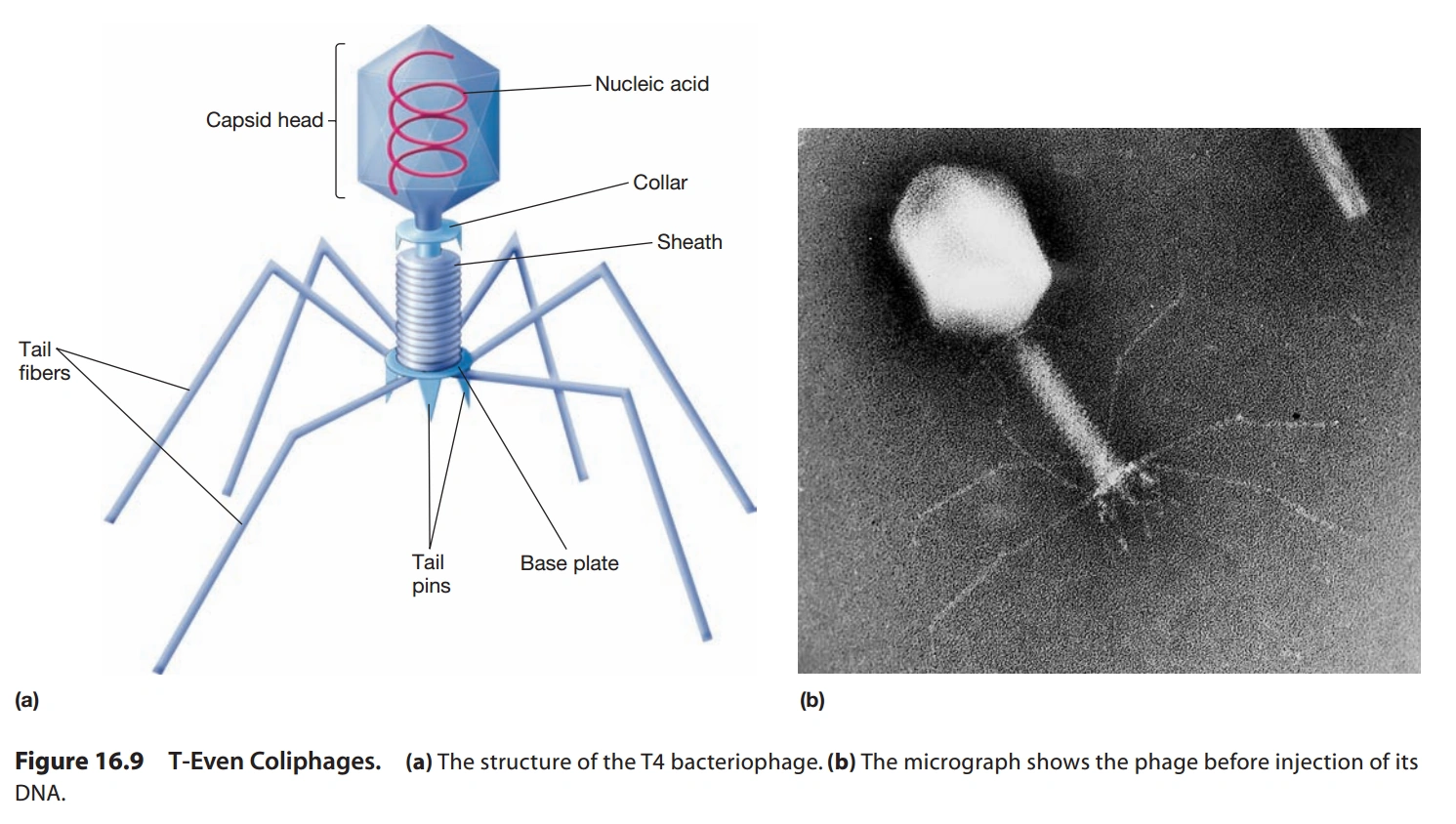
图5:T4噬菌体的结构

{kind=link}
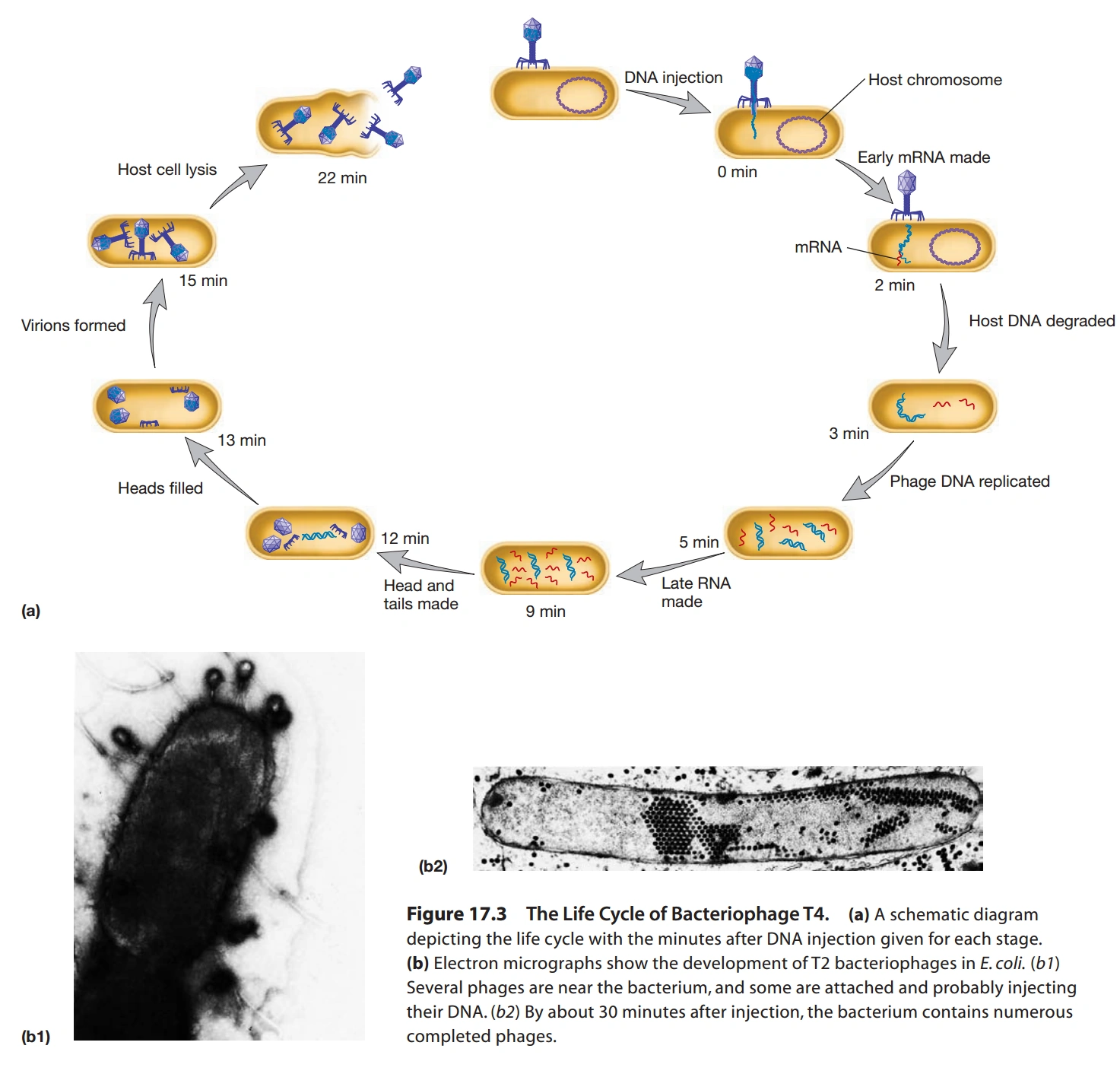
图6:T4噬菌体感染大肠杆菌的过程
{kind=link}
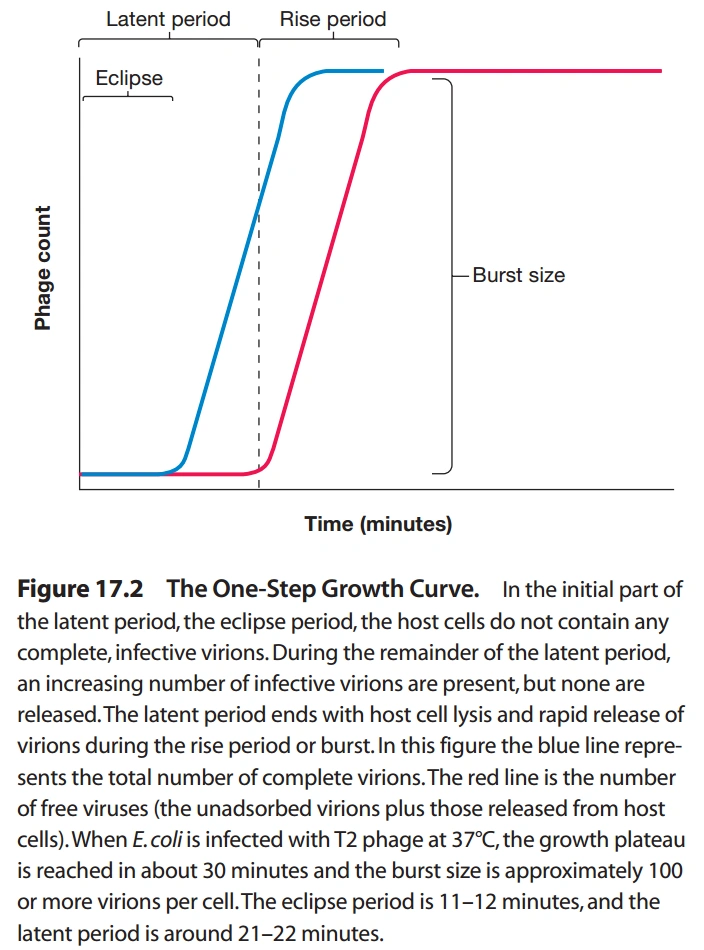
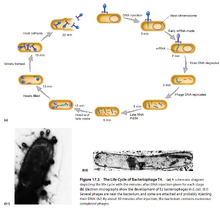
图7:T4噬菌体的生活史

{kind=link}
图8:盖病毒科核酸进入细菌的方式

{kind=link}
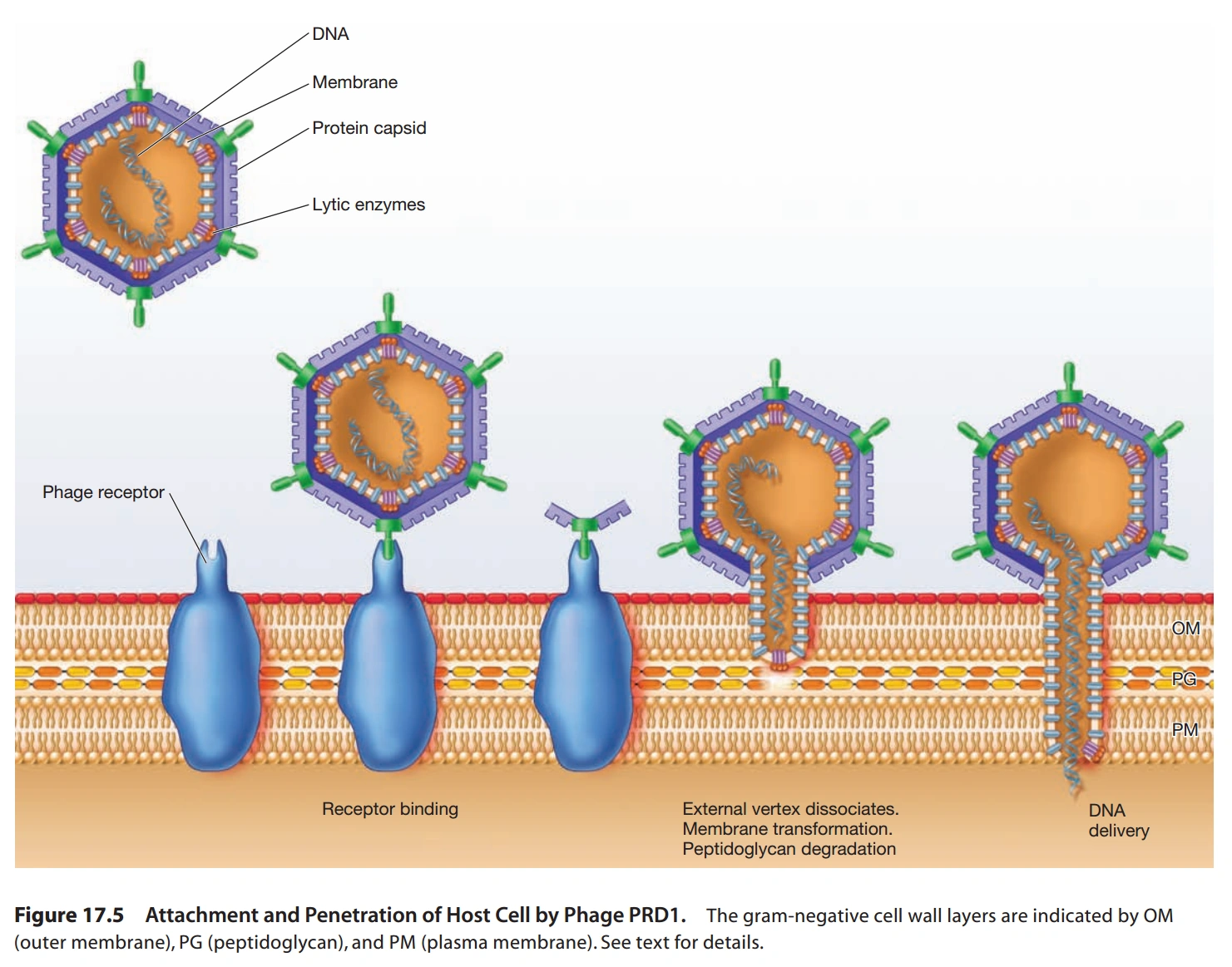
图9:T4噬菌体的DNA复制方式

{kind=link}
图10:其它噬菌体的结构

{kind=link}
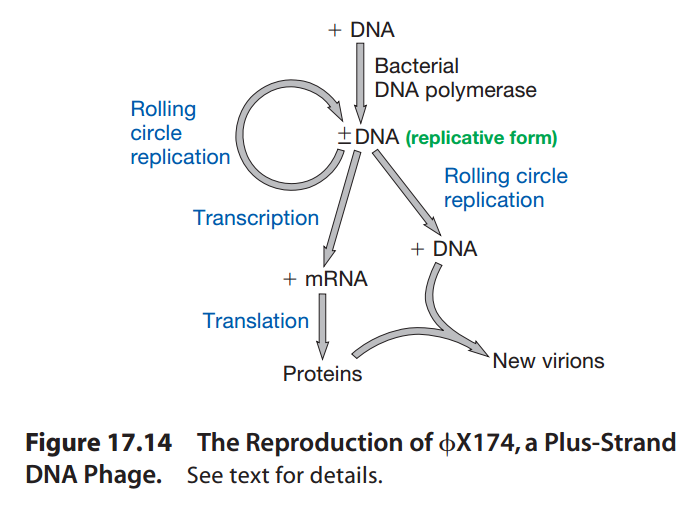
图11:φX174病毒的复制过程
{kind=link}
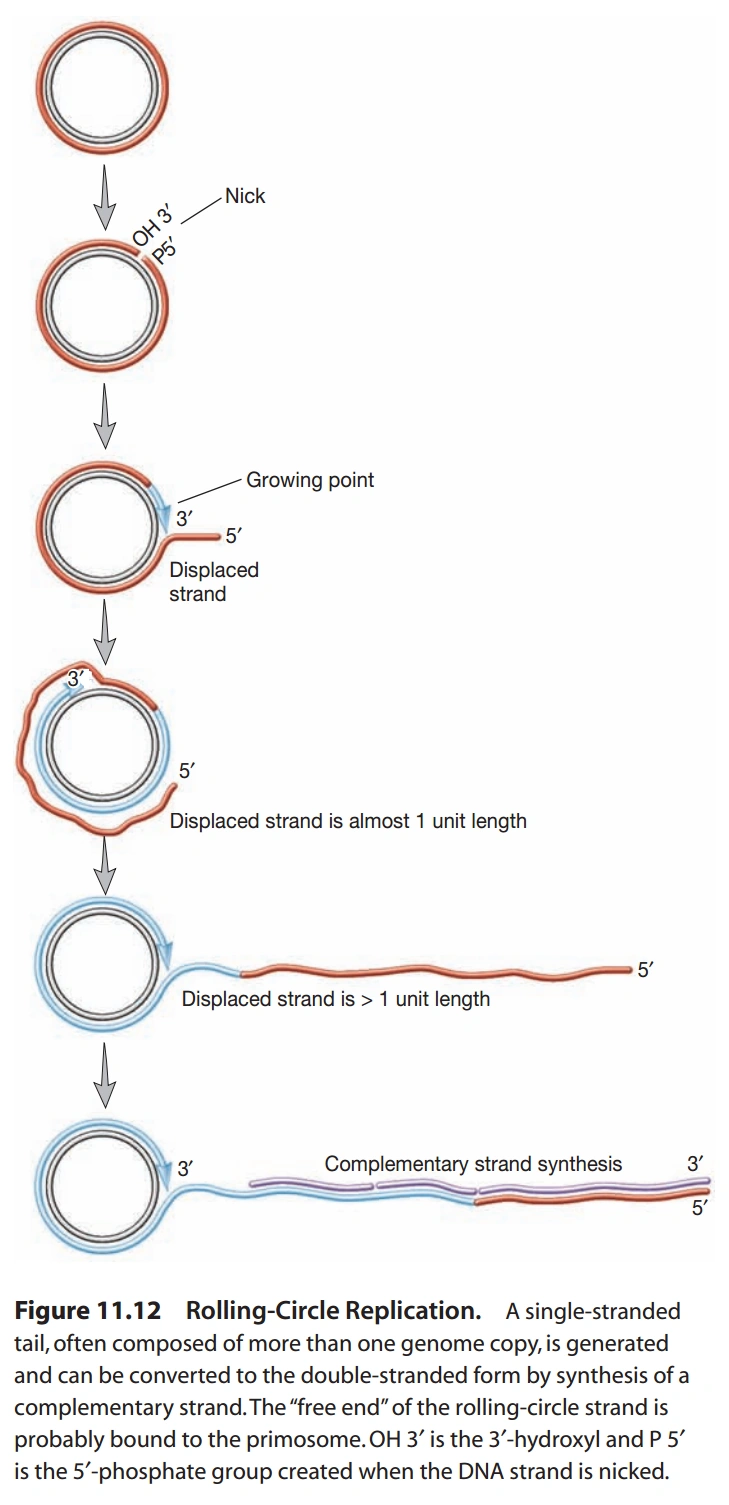
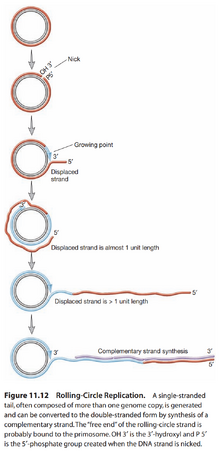
图12:滚轮式复制
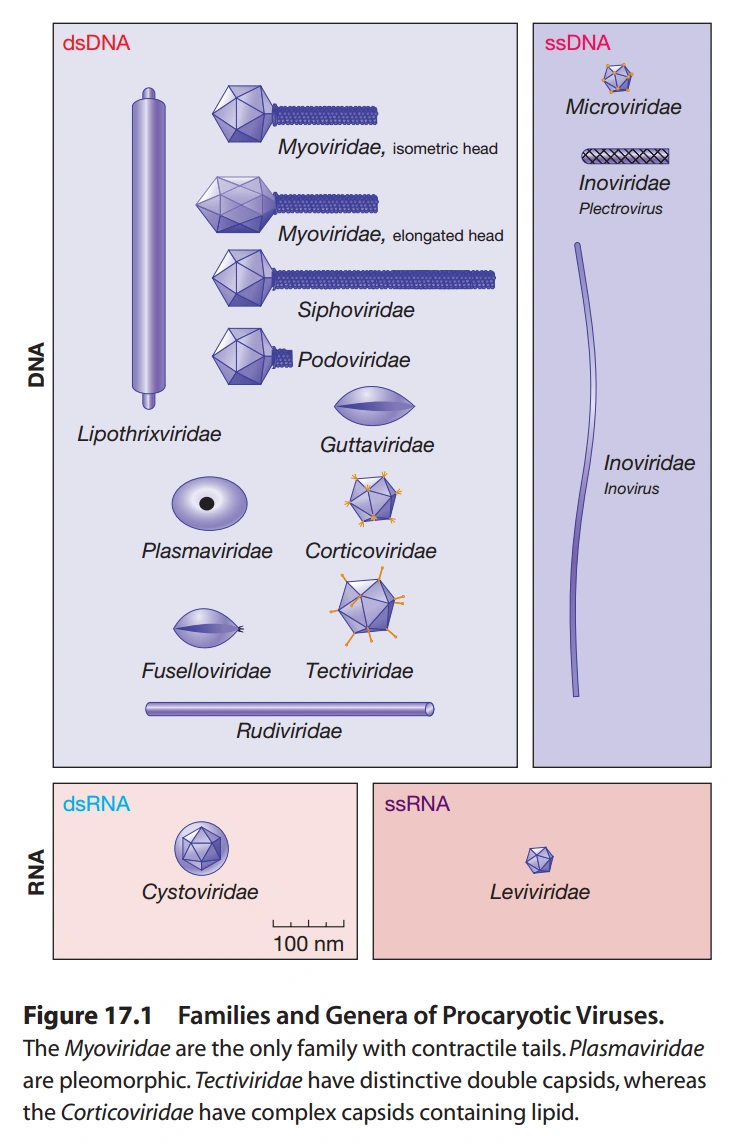
- 噬菌体(如未注明,一般感染大多数细菌,不感染古菌)
- 双链DNA病毒
- 肌病毒科(Myoviridae):
- 肌病毒科包含研究最多的噬菌体,即T4噬菌体和相关的T-偶噬菌体(T-even phage)。
- T4噬菌体一般感染革兰氏阴性菌。(其它肌病毒一般能感染大多数细菌,部分还能感染古菌)
- 具有正二十面体的头部、螺旋管状的尾部,尾部有纤维。(图5)
- 头部正二十面体可沿尾部的轴伸长(如T4)。
- 无脂双层膜,线形DNA,少数环形DNA。
- DNA中,胞嘧啶被羟甲胞嘧啶(HMC)替换,它可被糖基修饰。
- HMC的糖基防止病毒的DNA被细菌的限制性内切酶降解。
- 糖基HMC使病毒DNA与宿主DNA区分开,防止DNA被自己的酶破坏。(见下文)
- T4噬菌体感染大肠杆菌后,分为潜伏期(Latent Period)、上升期(Rise Period)、高原期(Plateau Period)。
- 潜伏期的前期,细胞内检测不到完整的病毒颗粒,称为隐藏期(Eclipse Period)。
- T4噬菌体的分子生活史:
- 首先,病毒的尾纤维和细菌表面的受体接触。
- 尾部的基盘随后和细菌表面接触。
- 病毒和细菌的结合受Mg2+、Ca2+、pH的影响。
- 然后,病毒的尾部变短,尾部核心蛋白插入细菌内。
- 核心蛋白在细胞膜上钻孔,允许DNA通过。
- DNA产生早期mRNA(Early mRNA),利用核糖体合成蛋白质,包括一些酶。
- T4噬菌体的酶会降解细胞核中原有的核酸,为合成病毒核酸提供原料。
- DNA的复制同时进行,病毒的DNA有多个复制原点。
- DNA复制完成后转录的mRNA称为晚期mRNA(Late mRNA)。
- 早期mRNA通常逆时针方向转录;晚期mRNA则顺时针方向转录。
- T4噬菌体利用核糖-ADP和RNA聚合酶的α亚基调控自身基因表达。
- 最后,T4产生穿孔素(Holin)和T4溶菌酶(T4 Lysozyme)。
- 穿孔素在细菌细胞膜上打孔,T4溶菌酶穿过孔攻击细菌的肽聚糖。
- 肽聚糖分解后,细菌破裂,释放病毒颗粒。
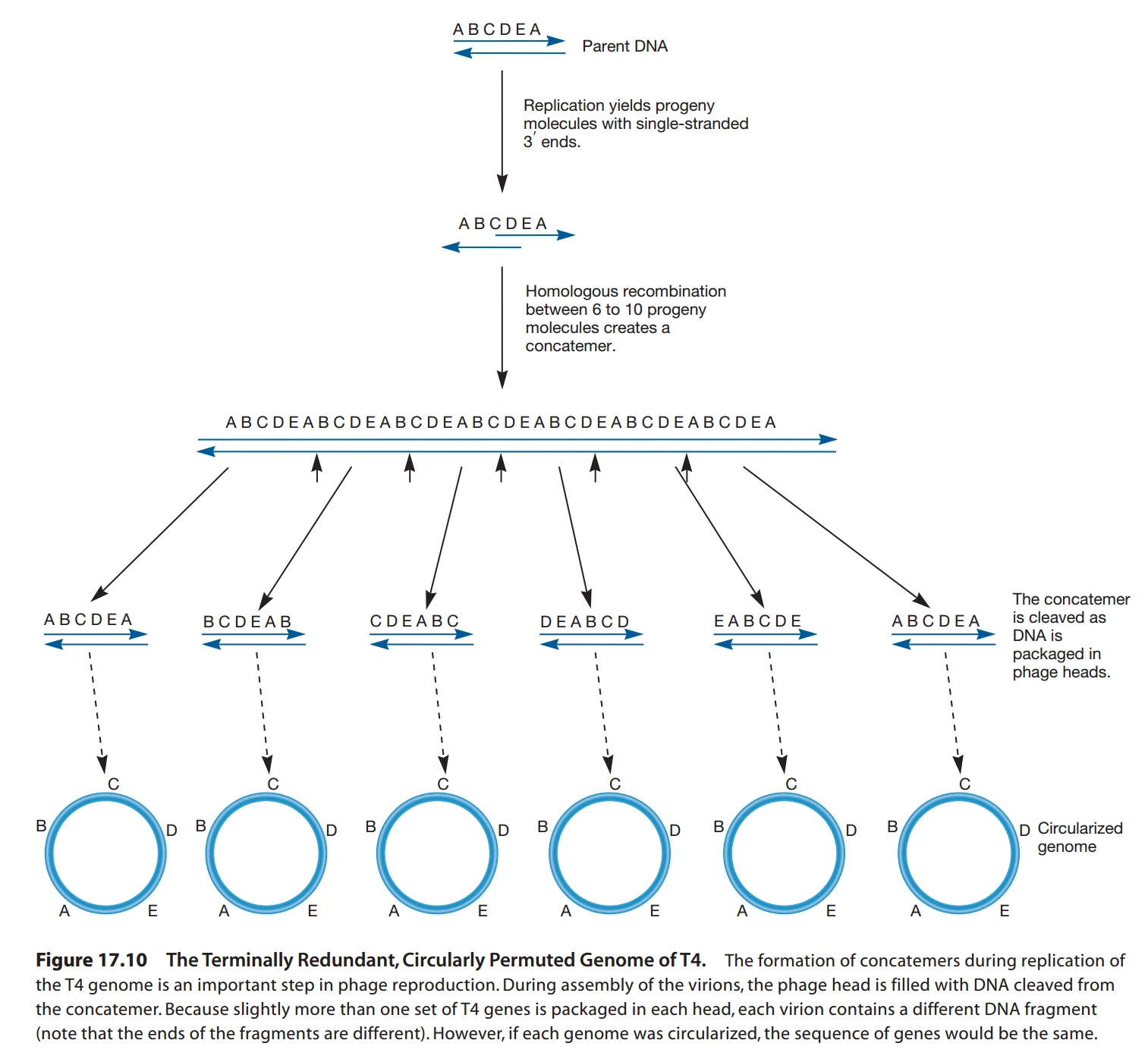
- 噬菌体DNA没有端粒,末端不能正常复制,T4噬菌体解决此问题的方法如下:
- DNA有多个复制原点,双向复制,末端不复制,形成末端部分单链的双链DNA。
- DNA两头有相似的末端冗余序列(Terminal Redundancy),基因组内部也有类似序列。
- 双向复制出的两条DNA杂交,形成串联体(Concatemer),随后被细菌的DNA连接酶连接。
- 一种病毒编码的限制性内切酶在特定序列部位切断串联体,形成新DNA。
- 切割的位置不一定是原来的连接部位,故切割结果好像是经过环形重组的。
- 因为有上述过程,T4噬菌体的基因组总是以环形绘制的。
- 若有多种类似T4噬菌体的病毒感染大肠杆菌,上述过程能使这些病毒交流基因,促进进化。
- 长尾噬菌体科(Siphoviridae):无脂双层膜,线形DNA,结构类似肌噬菌体科病毒,尾部较长,无末端冗余序列和环形重组现象,少数能感染细菌和古菌。
- 本科包含著名的λ噬菌体。
- λ噬菌体将DNA注入大肠杆菌后,它立刻环化形成类似质粒的结构。
- 之后可以有两种途径,一种是立刻大量复制产生新噬菌体,将大肠杆菌裂解。
- 另一种是将DNA整合入大肠杆菌的DNA,并随细菌复制而复制,这种细菌迟早被裂解,称为溶原性细菌(Lysogenic)。
- 短尾噬菌体科(Podoviridae):无脂双层膜,线形DNA,结构类似肌噬菌体科病毒,尾部较短,大部分无基因组环形重组,但有末端冗余序列,少部分有环形重组现象,少数只感染革兰氏阳性菌。
- 脂毛噬菌体科(Lipothrixviridae):杆状,有脂双层膜,线形DNA,DNA末端结构未知,感染古菌。
- 滴状病毒科(Guttaviridae):椭球状,有脂双层膜,环形DNA,DNA高度甲基化,感染古菌。
- 原生病毒科(Plasmaviridae):球状,有脂双层膜,环形DNA,感染支原体。
- 被脂病毒科(Corticoviridae):正二十面体,无膜包被,但外壳内有脂双层膜,环形DNA,DNA高度卷曲,但有较多负超螺旋。
- 微小纺锤体形病毒科(Fuselloviridae):柠檬形,有脂双层膜,环形DNA,DNA高度卷曲,病毒自带DNA整合酶,将自身DNA整合至宿主染色体的一个tRNA基因中,感染嗜热古菌。
- 盖病毒科(Tectiviridae):正二十面体,无脂双层膜,线形DNA,只感染革兰氏阴性菌。它是第二种核酸进入细胞的方式研究较多的病毒,见图8。
- 古病毒科(Rudiviridae):杆状,其中一端有三条纤维,无脂双层膜,线形DNA,DNA末端有冗余序列,DNA两条链共价相连,感染嗜热古菌。
- 总结:
- 肌病毒科、长尾噬菌体科、短尾噬菌体科共同构成有尾噬菌体目(Caudovirales),其中只有肌病毒科的尾部能收缩。
- 脂毛噬菌体科、古病毒科共同构成Ligamenvirales目。
- 具有正二十面体结构的病毒都无膜包被,都是线形DNA,除被脂病毒科外都无脂双层膜。
- 不具有正二十面体结构的病毒都有膜包被,古病毒科除外,除古病毒科和脂毛噬菌体科外都是环形DNA。
- 能感染古菌的病毒有:肌病毒科、长尾噬菌体科、脂毛噬菌体科、滴状病毒科、微小纺锤体形病毒科、古病毒科。
- 只感染革兰氏阴性菌的病毒有:肌病毒科(部分,特别是T4噬菌体)、原生病毒科(如果把支原体算作革兰氏阴性菌)。
- 只感染革兰氏阳性菌的病毒有:短尾噬菌体科(少数)。
- 有DNA环形重组现象的科:肌病毒科、短尾噬菌体科。
- 肌病毒科(Myoviridae):
- 单链DNA病毒:以下两科病毒DNA复制方式特殊,称为滚轮式复制(Rolling Circle Replication),见图12。
- 微小噬菌体科(Microviridae):
- 本科包含研究较多的φX174病毒。
- 正二十面体,无脂双层膜,环形DNA。
- 本科病毒感染:蛭弧菌(Bdellovibrio)、肠杆菌(Enterobacteria)、衣原体(Chlamidia)、螺原菌(Spiroplasma)。(除螺原菌外,都是革兰氏阴性菌)
- φX174病毒的核酸是正链。
- 核酸进入细胞后,利用细菌的DNA聚合酶从单链DNA合成双链DNA,称为复制形态(Replicative Form)。
- 复制形态的DNA指导细菌生产更多的复制形态DNA(指导蛋白质合成),然后生产单链DNA(用于组装病毒)。
- 病毒组装完成后,病毒产生“酶E”(Enzyme E),抑制细菌的肽聚糖合成。
- 细胞壁因缺少肽聚糖变弱,细菌裂解,释放病毒颗粒。
- 丝状噬菌体科(Inoviridae):
- 本科包含研究较多的fd噬菌体。
- 杆状,无脂双层膜,环形DNA,部分只感染革兰氏阴性菌。
- fd噬菌体只感染有F因子(F+、Hfr、F')的大肠杆菌。
- fd噬菌体通过附着在F因子编码的鞭毛上感染大肠杆菌。
- fd噬菌体复制过程类似φX174病毒。
- fd噬菌体不会杀死宿主,而是使宿主将病毒颗粒分泌到胞外。
- 微小噬菌体科(Microviridae):
- RNA病毒
- 双链RNA病毒:囊状噬菌体科(Cystoviridae),球形,有脂双层膜,线形DNA,DNA分为三段(L、M、S),每段编码一个mRNA,翻译为多种蛋白质。有双层蛋白质外壳,呈略不同的正二十面体。只感染假单胞菌(Pseudomonas),尤其是丁香假单胞菌(P. syringae)。
- 单链正链RNA病毒:光滑病毒科(Leviviridae):
- 本科包含研究较多的MS2和Qβ噬菌体。
- 正二十面体,无脂双层膜,线形DNA,DNA 5’端有甲基化G帽。
- 它们感染有F因子的大肠杆菌。
- 它们利用大肠杆菌表面的F鞭毛将核酸插入细胞。
- 进入细胞的核酸直接作为mRNA指导蛋白质合成。
- 最先合成的蛋白质是RNA复制酶(RNA Replicase)。
- RNA复制酶将RNA补为双链RNA,称为复制形态。
- RNA复制酶利用复制形态的RNA生成大量单链RNA。
- 一部分单链RNA被用于合成RNA,其它被用于指导蛋白质合成。
- 病毒裂解细胞的方式类似φX174病毒。
- 双链DNA病毒
- 真核生物病毒(以人类的病毒为重点)
- DNA病毒
- 疱疹病毒科(Herpesviridae):
- 本科包含HSV病毒(Herpes Simplex Virus,单纯疱疹病毒,分2种)、VZV病毒(Varicella Zoster Virus,水痘-带状疱疹病毒)、EB病毒(Epstein-Barr Virus)、巨细胞病毒(Cytomegalovirus)、HHV6、HHV7、卡波希氏肉瘤病毒(Kaposi's Sarcoma-associated Herpesvirus)。(依次称为人类疱疹病毒1-8号)
- 不定形,有脂双层膜,线形双链DNA,DNA末端和内部有重复序列。
- 病毒表面的糖蛋白与细胞表面的硫酸乙酰肝素类受体(Heparan Sulfate-like Receptor)、TNF类受体、免疫球蛋白类受体结合。
- 脂双层膜与细胞融合,蛋白质外壳及核酸进入细胞质,释放若干种蛋白质。
- 蛋白质外壳在细胞核膜处钻孔,释放DNA,DNA进入细胞核后立即环化。
- HSV的基因分为三类(记为α、β、γ)。
- α基因编码的蛋白质为DNA复制做准备。
- β基因编码的蛋白质主要为DNA复制所需的酶和调控蛋白。
- γ基因必须在DNA复制后表达,编码大多数结构蛋白。
- DNA首先用类似细菌环形DNA复制的方式复制。
- 然后DNA用滚轮式复制产生串联体。
- 串联体切割产生病毒DNA,无环形重组现象。
- 病毒组装在细胞核进行,病毒离开细胞核内膜时获得脂双层膜。
- 完整的病毒通过内质网和高尔基体利用胞吐离开细胞。
- HSV病毒感染部分神经细胞(特别是神经节细胞)时,基因不表达,称为潜伏期(Latency)。(其它疱疹病毒也有潜伏期,但感染部位不是神经细胞而是与免疫系统有关的细胞)
- 潜伏期的病毒会表达LAT RNA,它调控宿主基因的表达,干扰宿主细胞凋亡。
- 潜伏期病毒的基因组可能嵌入神经细胞的染色体。
- 嵌入神经细胞染色体的病毒基因组会造成感染症状周期性爆发。
- 肝病毒科(Hepadnaviridae):
- 本科包含乙肝病毒(Hepatitis B Virus,HBV)。
- 蛋白质外壳呈正二十面体,有脂双层膜,环形DNA,DNA既有双链也有单链部分,外壳内有DNA聚合酶。
- 乙肝病毒的DNA的两条链都是线形的;负链是完整的;正链是不完整的,在负链两端的位置却是相连的,所以整个DNA是环形的。
- 乙肝病毒复制过程存在反转录过程,但不属于反转录病毒。(称为副反转录病毒(Pararetrovirus))
- 乙肝病毒DNA有时会通过重组进入宿主染色体,但此时病毒基因失活。
- 进入宿主染色体的病毒DNA会促进细胞分裂,可能引发肝癌。
- 病毒进入细胞的过程类似疱疹病毒。
- DNA进入细胞核后,被修复酶补成完整的环形DNA(cccDNA)。(两条链都是环形的)
- cccDNA有来自宿主的组蛋白,称为微染色体(Minichromosome)。
- DNA转录出若干mRNA,其中一种称为预基因组(Pregenome)。
- 一种mRNA转录出的P蛋白有三种酶的活性:RNA反转录酶、DNA聚合酶、RNA水解酶。
- P蛋白和预基因组、结构蛋白结合形成不成熟的病毒颗粒。
- 病毒颗粒内,P蛋白反转录预基因组,形成DNA的负链,然后从负链合成DNA的正链。
- 成熟的病毒颗粒既可运回细胞核增殖,也可在内质网处获得脂双层膜,分泌出细胞。
- 乙肝病毒是基因组最小的DNA病毒。
- 细小病毒科(Parvovirus):
- 本科包含B19病毒。
- 正二十面体,无脂双层膜,线形单链DNA。
- 细小病毒的基因组非常简单,只编码三个蛋白质,都是结构蛋白。
- 细小病毒必须完全依赖宿主的酶复制,因而只能在细胞周期的S期增殖。
- 细小病毒DNA的末端与内部的一段形成互补双链,作为DNA复制的引物,欺骗DNA聚合酶复制病毒DNA。
- 乳头瘤病毒科(Papillomaviridae):
- 本科包含HPV病毒(Human Papillomavirus,人类乳头瘤病毒)。
- 球状,无脂双层膜,环形双链DNA,DNA有组蛋白。
- 人类乳头瘤病毒感染人的鳞状上皮组织(Squamous Epithelium)的基部细胞(Basal Cells)。
- 之后,病毒随着上皮细胞的分化进入角化细胞(Keratinocyte)。
- 病毒编码的蛋白质会降解细胞中的p53和Rb蛋白,引起癌变。
- 类似于乙肝病毒,HPV的DNA也会通过基因重组进入宿主染色体,引发癌变。
- 细胞上与病毒结合的受体可能是硫酸乙酰肝素受体,但有争议。
- 病毒通过胞吞进入细胞。
- 细胞内的还原性环境破坏病毒外壳上的二硫键,释放DNA,DNA进入细胞核。
- DNA随上皮细胞的分裂而复制,直到上皮细胞分化为角化细胞。
- 角化细胞中,DNA大量复制,同时表达结构蛋白,病毒组装在细胞核中进行。
- 多瘤病毒科(Polyomaviridae):
- 本科包含JC病毒、BK病毒、默克尔细胞多瘤病毒(Merkel Cell Polyoma Virus)。
- 结构与乳头瘤病毒相似。
- DNA复制在细胞核中进行,病毒组装在细胞核中的病毒工厂进行,生活史很普通。
- 病毒通过裂解细胞释放。
- 腺病毒科(Adenoviridae):
- 本科包含人腺病毒(Human Adenovirus)。
- 球状和正二十面体状之间,无脂双层膜,线形双链DNA,两个末端有互补的冗余序列,末端与蛋白质共价连接。
- DNA复制、病毒组装在细胞核中进行。
- DNA复制使用特殊的链取代法(Strand Replacement)。
- 首先,模板DNA从末端解开,根据其中一条链合成与它互补的链,取代原有的另一条链。
- 被取代的链立刻与特殊蛋白结合,防止它与其它模板DNA杂交。
- 被取代的链因两个末端有互补序列而自身杂交变为环状。
- 同样的DNA聚合酶把被取代的链补为双链DNA。
- 链取代法的特征是每次只合成一条DNA链。
- 痘病毒科(Poxviridae):
- 本科包含天花病毒(Variola Virus)、口疮病毒(Orf Virus)、接触传染性软疣病毒(Molluscum Contagiosum Virus)、牛痘病毒(Vaccinia Virus)。
- 椭球状或砖块状,有脂双层膜,蛋白质外壳呈杆状但可弯曲,线形双链DNA,在两个末端两条链共价相连。
- 病毒自带RNA和DNA聚合酶,在细胞质病毒工厂中转录和复制DNA、组装病毒。
- DNA复制也以链取代法进行。
- 病毒颗粒既可通过裂解也可通过分泌离开细胞。
- 疱疹病毒科(Herpesviridae):
- 单链正链RNA病毒
- 微小RNA病毒科(Picornaviridae):
- 本科包含脊髓灰质炎病毒(Poliovirus)、柯萨基病毒(Coxsackievirus)、埃可病毒(Echovirus)、EV病毒(Enterovirus)、鼻病毒(Rhinovirus)、甲肝病毒(Hepatitis A Virus)。
- 球状,无脂双层膜,线形RNA,整个RNA结构类似一个mRNA,但无5‘端G帽,而是共价连接一个蛋白质,5’端有三叶草形IRES结构,是核糖体开始翻译的部位。
- 病毒通过胞吞进入细胞。
- 内体中,外壳蛋白构象改变,释放VP4蛋白,在内体膜上开孔。
- RNA被释放入细胞质,5‘端的蛋白被去除,然后整个RNA作为一个mRNA被翻译。
- 翻译出的蛋白质经切割成为多种蛋白。
- 病毒的蛋白酶切割翻译的启动因子,阻止正常的(有5‘端G帽的)mRNA翻译。
- RNA复制在由内质网膜产生的有两层膜的囊泡(病毒工厂)中进行。
- 囊泡内,RNA先被补全为双链RNA,然后以类似链取代法复制。(但不能称作“链取代法”)
- 病毒通过裂解细胞释放。
- 杯状病毒科(Caliciviridae):
- 本科包含诺洛病毒(Norovirus)、戊肝病毒(Hepatitis E Virus)。
- 正二十面体,无脂双层膜,线形RNA,RNA 5‘端共价连接有蛋白质,3’端有多聚A尾巴。
- 生活史类似微小RNA病毒科。
- 病毒通过裂解细胞释放。
- 披盖病毒科(Togaviridae):
- 本科包含风疹病毒(Rubella Virus)、(东部、西部)马脑炎病毒(Eastern/Western Equine Encephalitis Virus)。
- 球状,有脂双层膜,蛋白质外壳呈正二十面体,线形RNA,RNA 5‘端有G帽,3’端有多聚A尾巴。
- 生活史类似微小RNA病毒科。
- 黄病毒科(Flaviviridae):
- 本科包含黄热病毒(Yellow Fever Virus)、登革热病毒(Dengue Virus)、圣路易斯脑炎病毒(St. Louis Encephalitis Virus)、西尼罗病毒(West Nile Virus)、丙肝病毒(Hepatitis C Virus)、庚肝病毒(Hepatitis G Virus)。
- 注:尚无证据证明庚肝病毒能引起或参与肝炎,事实上目前没有确认庚肝引起任何人类疾病。有临床数据显示HIV感染者,若又感染HGV,可比单独感染HIV者活得更长。
- 球状,有脂双层膜,线形RNA,RNA 5‘端有G帽或共价连接有蛋白质,3’端无多聚A尾巴,但有环形结构。
- 生活史类似微小RNA病毒科。
- 冠状病毒科(Coronaviridae):
- 本科包含SARS病毒(Severe Acute Respiratory Syndrome,严重急性呼吸道综合症)和MERS病毒(Middle East Respiratory Syndrome,中东呼吸综合症)。
- 球状,有脂双层膜,线形RNA,RNA与N蛋白共价相连,N蛋白参与蛋白质外壳。
- 冠状病毒的RNA是RNA病毒中最大的。
- 生活史类似微小RNA病毒科。
- 微小RNA病毒科(Picornaviridae):
- 单链负链RNA病毒
- 棒状病毒科(Rhabdoviridae):
- 本科包含狂犬病病毒(Rabies Virus)、水疱性口炎病毒(Vesicular Stomatitis Virus)。
- 子弹形,线形RNA,有脂双层膜,RNA共价结合有N蛋白。
- 病毒自带RNA聚合酶,在细胞质中转录RNA。
- RNA聚合酶先在3‘端解链,转录一小段(称为Leader RNA),然后终止,再在RNA上寻找转录起始信号(Transcription Start Signal),转录至转录终止信号(Transcription Stop Signal)。
- 转录出的RNA(Leader RNA除外)5’端有G帽,3‘端有多聚A尾巴。
- 模板DNA上只有少量U碱基,RNA聚合酶遇到转录终止信号时,每遇到U碱基,在加上一个A碱基时会往后退一格,这样少量的U会被转录很多次,从而形成多聚A尾巴。(称为Shuttering)
- Leader RNA和模板RNA杂交,然后被RNA聚合酶补全为正链RNA,最后正链RNA被转录,形成新的负链RNA。
- 病毒组装在细胞质中进行。
- 纤丝病毒科(Filoviridae):
- 本科包含马尔堡病毒(Marburg Virus)、埃博拉病毒(Ebola Virus)。
- 蛇状,有脂双层膜,线形RNA,DNA共价结合有N蛋白。
- 生活史类似棒状病毒科。
- 副粘病毒科(Paramyxoviridae):
- 本科包含副流感病毒(Parainfluenza Virus)、呼吸道合胞体病毒(Respiratory Syncytial Virus)、新城鸡瘟病毒(Newcastle Disease Virus)、腮腺炎病毒(Mumps Virus)、麻疹病毒(Rubeola Virus)。
- 球状,有脂双层膜,膜上有血凝素和神经氨酸苷酶,线形RNA,RNA与N蛋白共价相连。
- 生活史类似棒状病毒科。
- 正黏病毒科(Orthomyxoviridae):
- 本科包含(甲、乙、丙型)流感病毒(Influenza A/B/C)。
- 结构类似副粘病毒科,RNA分为8段,RNA共价结合有N蛋白。
- 生活史类似棒状病毒科。
- 甲型流感病毒根据其含的血凝素(Hemagglutinin)和神经氨酸苷酶(Neuraminidase)的不同,分为18(H)×11(N)= 198种,其中常见的引起人类疾病的有H1、H2、H3、N1、N2。(乙、丙型流感病毒也有血凝素和神经氨酸苷酶,但变异不大,故不分类)
- 历次流感大爆发:H3N8或H2N2(1889-1890)、H1N1(1918-1920)、H2N2(1957-1958)H3N2(1968-1969)、H1N1(1977-1978)、H1N1或H1N9(2009-2010)。
- 布尼亚病毒科(Bunyaviridae):
- 本科包含汉坦病毒(Hantavirus)、加州脑炎病毒(California Encephalitis Virus)、白蛉热病毒(Sandfly Fever Virus)。
- 球状,有脂双层膜,RNA分三段,2段环形,1段线形,RNA共价结合有N蛋白。
- 生活史类似棒状病毒科。
- 砂粒病毒科(Arenaviridae):
- 本科包含淋巴细胞性脉络丛脑膜炎病毒(Lymphocytic Choriomeningitis Virus)、拉沙热病毒(Lassa Fever Virus)、南美出血热病毒(South American Hemorrhagic Fever Virus)。
- 球状,有脂双层膜,RNA分两段,线形RNA,RNA共价结合有N蛋白。
- 生活史类似棒状病毒科。
- 丁肝病毒属(Deltavirus):
- 本属不属于任何科,只包含丁肝病毒。
- 丁肝病毒属于缺陷病毒,只能和乙肝同时感染或感染已携带乙肝病毒者。
- 球状,有脂双层膜,环形RNA,RNA复制采用滚轮式复制。
- 丁肝自带核酶,用于修饰核酸。
- 棒状病毒科(Rhabdoviridae):
- 双链RNA病毒
- 呼肠孤病毒科(Reoviridae):
- 本科包含轮状病毒(Rotavirus)、呼肠病毒(Reovirus)、科罗拉多蜱热病毒(Colorado Tick Fever Virus)。
- 球状,无脂双层膜,有两层蛋白质外壳(部分只有一层),线形RNA,RNA分10-12段。
- RNA 5'端可有G帽,也可有蛋白质,亦可什么都没有。
- 病毒在生活史中从不完全脱去蛋白质外壳,因而不会引起普通免疫反应。
- 病毒自带的RNA聚合酶完成转录和复制。
- 呼肠孤病毒科(Reoviridae):
- 反转录病毒
- 反转录病毒科(Retroviridae)
- 本科病毒又分7类,称为Alpharetrovirus、Betaretrovirus、Gammaretrovirus、Deltaretrovirus、Epsilonretrovirus、Lentivirus、Spumavirus。
- Alpharetrovirus包含劳氏肉瘤病毒(Rous Sarcoma Virus),它是最小的RNA病毒。
- Betaretrovirus包含小鼠乳房肉瘤病毒(Mouse Mammary Tumor Virus)。
- Gammaretrovirus包含鼠白血病病毒(Murine Leukemia Virus)。
- Deltaretrovirus包含人类嗜T细胞病毒(Human T Lymphotropic Virus)和猴类嗜T细胞病毒(Simian T Lymphotropic Virus)。
- Epsilonretrovirus包含大眼梭鲈皮肤肉瘤病毒(Walleye Dermal Sarcoma Virus)。
- Lentivirus包含人类免疫缺陷病毒(Human Immunodeficiency Virus)。
- Spumavirus包含猴泡沫病毒(Simian Foamy Virus)。
- Epsilonretrovirus和Spumavirus不含人类病原体。
- 有些反转录病毒感染远古脊椎动物后变为基因组的一部分(常为转位子),特定条件下它会被激活,称为内源性反转录病毒(Endogenous),如小鼠乳房肉瘤病毒。
- 球状,有脂双层膜,线形单链RNA,有两份,RNA 5’端有G帽,3‘端有尾巴,自带反转录酶和整合酶。
- 病毒通过胞吞进入细胞,反转录酶通过单链RNA合成双链DNA。
- 双链DNA被整合酶随机整合入宿主基因组。
- RNA聚合酶II转录病毒DNA,一部分被用于翻译,一部分被用于组装病毒。
- 反转录病毒科(Retroviridae)
- 总结和补充:
- DNA病毒只有一半有脂双层膜(疱疹病毒科、肝病毒科、痘病毒科)。
- RNA病毒大多有脂双层膜,少数除外(微小RNA病毒科、杯状病毒科、呼肠孤病毒科)。
- DNA病毒大多是双链病毒,细小病毒科除外;RNA病毒大多是单链病毒,呼肠孤病毒科除外。
- RNA大多是线形,布尼亚病毒科和丁肝病毒属除外。
- DNA一般是线形,包括疱疹病毒科、细小病毒科、腺病毒科、痘病毒科。
- DNA病毒的核酸合成部位一般在细胞核,肝病毒科和痘病毒科除外。
- RNA病毒的核酸合成部位一般在细胞质,正黏病毒科除外,反转录病毒一部分在细胞核。
- 脂双层膜来源:肝病毒科的来自内质网,疱疹病毒科的来自核膜,冠状病毒科的来自内质网和高尔基体,其它的来自细胞膜。
- 能引起感冒的病毒:鼻病毒(微小RNA病毒科)、肠病毒(微小RNA病毒科)、SARS和MERS(冠状病毒科)、人呼吸道合胞病毒(副粘病毒科)、副流感病毒(副粘病毒科)、人类偏肺病毒(副粘病毒科)、流感病毒(正粘病毒科)、腺病毒科、单纯疱疹病毒(疱疹病毒科)。
- DNA病毒
类似病毒的结构
- 朊病毒(Prion)是最小的感染性颗粒,它只有蛋白质成分,目前被认为是折叠错误的蛋白,它会误导更多的同类蛋白折叠错误,引起疾病。
- 只在哺乳动物和真菌中发现了朊病毒。
- 哺乳动物的朊病毒主要是PrP蛋白,α突触核蛋白(Alpha-synuclein)可能也是。
- PrPSc是PrP蛋白的错误形式,引起的疾病有疯牛病(牛海绵状脑疾病,Bovine Spongiform Encephalopathy)、羊瘙痒病(Scrapie)、CJD疾病、GSS综合征、家族性致命性失眠、库鲁病。
- α突触核蛋白被认为引起多系统萎缩疾病(MSA)。
- 折叠错误的蛋白在细胞中形成丝状的类淀粉蛋白(Amyloid),它是由大量β折叠片构成的,两端都可生长。
- PrP蛋白在细胞中的正常功能尚不明确,其结构中有大量α螺旋,它能与铜离子紧密结合,它一般出现在细胞膜上,既可跨膜又可通过糖脂(GPI)相连。
- 有人认为PrP参与细胞连接、细胞通讯、长期记忆形成、周围神经系统的髓鞘维持、骨髓造血干细胞的维持。
- 类病毒(Viroid)是第二小的感染性颗粒,只由RNA组成。
- 类病毒大多是植物的病原体。
- 类病毒被认为是远古的RNA世界的遗迹。
- 能被类病毒感染的物种:土豆、柠檬、啤酒花、葡萄藤、椰子、苹果、鸭梨、桃子、茄子、彩叶草(锦紫苏)。
- 细胞中,类病毒在细胞核或叶绿体中复制,一般需要RNA聚合酶II,复制机制为滚轮式复制。
- 有些类病毒是核酶。
- 一般的类病毒并不编码任何蛋白质,其致病机制一般认为是通过siRNA干扰植物正常mRNA。
- 拟病毒(Virusoid):环状单链RNA,必须和其它植物病毒同时感染,RNA只有几百个碱基,不编码任何蛋白质,离开植物细胞时有来自植物病毒的蛋白质包被。
- 噬病毒体(Virophage):线形双链DNA,必须和其它病毒同时感染,这一病毒一般是巨型病毒,噬病毒体自己有DNA聚合酶,但要依赖巨型病毒的RNA聚合酶转录,噬病毒体一般会感染巨型病毒的复制。
- 缺陷感染颗粒(Defective Interfering Particle):正常的病毒缺失了基因组的一段而无法独立完成在宿主细胞中复制,必须要和正常病毒同时感染才能复制。(几乎每一种DNA和RNA病毒都会自发产生DIP)
- 转位子(Transposable Genetic Element或Transposon):
- 转位子是可以在基因组中变换位置的基因片段,被称为“自私的基因”。
- 细菌、植物、酵母、动物基因中都存在大量转位子,人类基因组的45%都是转位子的碎片。
- 根据转位子变换位置的方式,将转位子分为三类,其中只有前两类常见:剪切式转位子、反转录式转位子、DNA复制式转位子。
- 细菌中最常见的转位子是IS元素,其特点是两端存在高度保守的回文序列,此序列变异会引起转位子无法移动。
- IS元素属于剪切型转位子,它引起的基因失活,一般会很快变异回到野生型。
- 两个IS元素位置很近时,它们可以带着中间的正常序列一起转位,这种现象称为复合转位子(Composite Transposon)。
- 细菌的Tn3转位子是DNA复制式转位子的代表,它首先将自身所在的DNA与一个环形质粒融合,两者相接的地方各有一个Tn3转位子,然后再将这个大质粒切断,产生的2个质粒各有一个Tn3转位子。
- 真核生物的转位子大部分是剪切式,最早发现的是玉米的Ac-Ds转位子。
- Ac和Ds是两个不同的转位子,Ac的序列几乎是固定的,它编码的是一个转位酶;Ds序列的两端与Ac相同,但内部很不相同,有些是Ac内部失去一部分序列形成的,有些是夹入正常基因组的一部分形成的,有些是Ds的内部又夹了一个Ds,其中第三类会引起染色体断裂。