{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
总论
- 刺胞动物门是动物界中第二原始的分类,约含11000个物种。(最原始的是海绵动物)
- 旧系统曾将刺胞动物门和栉水母动物门(Ctenophora)合并为腔肠动物门。
- 刺胞动物门尚未出现三胚层分化,但内胚层和外胚层间有来自外胚层的间质(Mesenchyme,有细胞)或中胶层(Mesoglea,无细胞),没有高等动物的基膜。
- 体型仍为辐射对称(Radial Symmetry),主轴为口腔反口轴(Oral-Aboral),无器官分化,甚至肌肉细胞和上皮细胞也未分化,称为肌皮细胞(Myoepithelial Cell)。
- 部分物种有世代交替现象,分水螅型(Polypoid)和水母型(Medusoid)。
- 没有体腔,只有内胚层围成的腔肠(Coelenteron),只有一个对外开口,既是嘴也是肛门。
- 出现了最原始的神经系统,呈网状。
- 幼虫称为浮浪幼虫(Planula),有鞭毛,能运动,有原肠胚。
- 主要分为四纲:水螅纲(Hydrozoa)、珊瑚纲(Anthozoa)、立方水母纲(Cubozoa)、钵水母纲(Scyphozoa)。
形态学
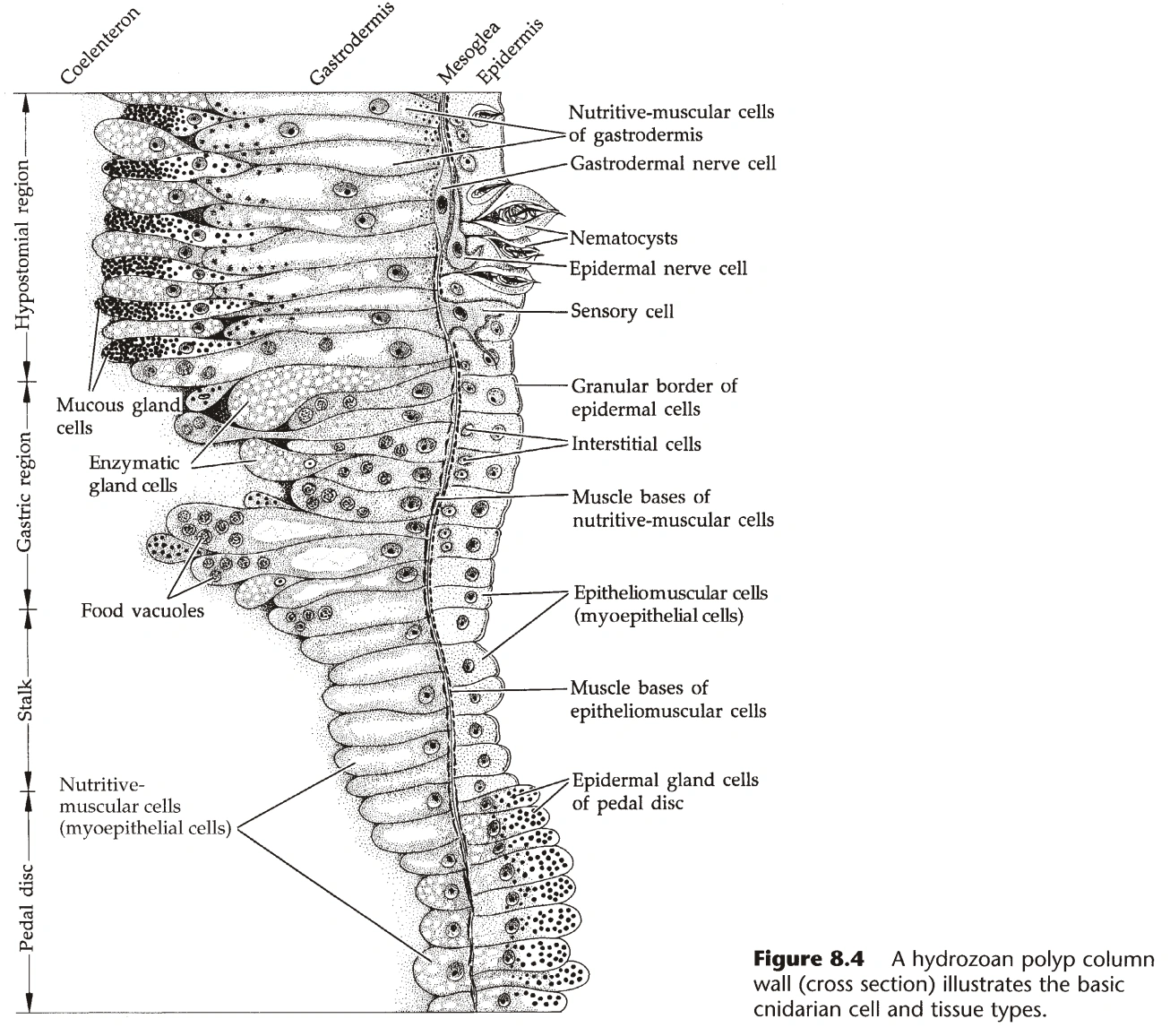
{kind=link}
图1:刺胞动物的基本组织结构
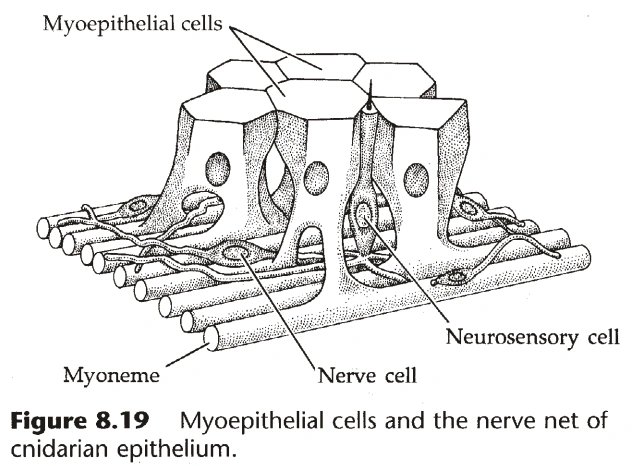
{kind=link}
图2:肌皮细胞
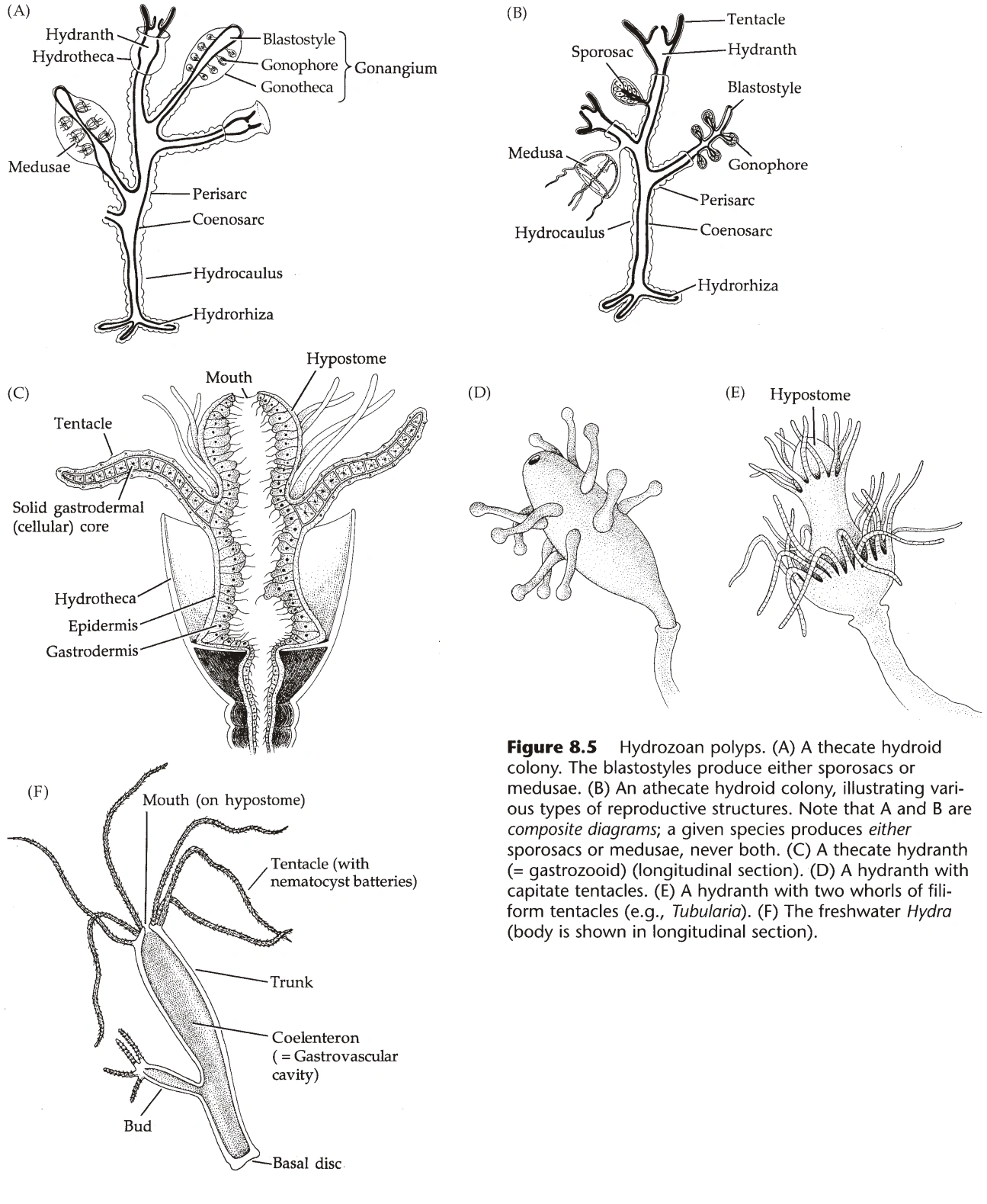
{kind=link}
图3:水螅纲的水螅型
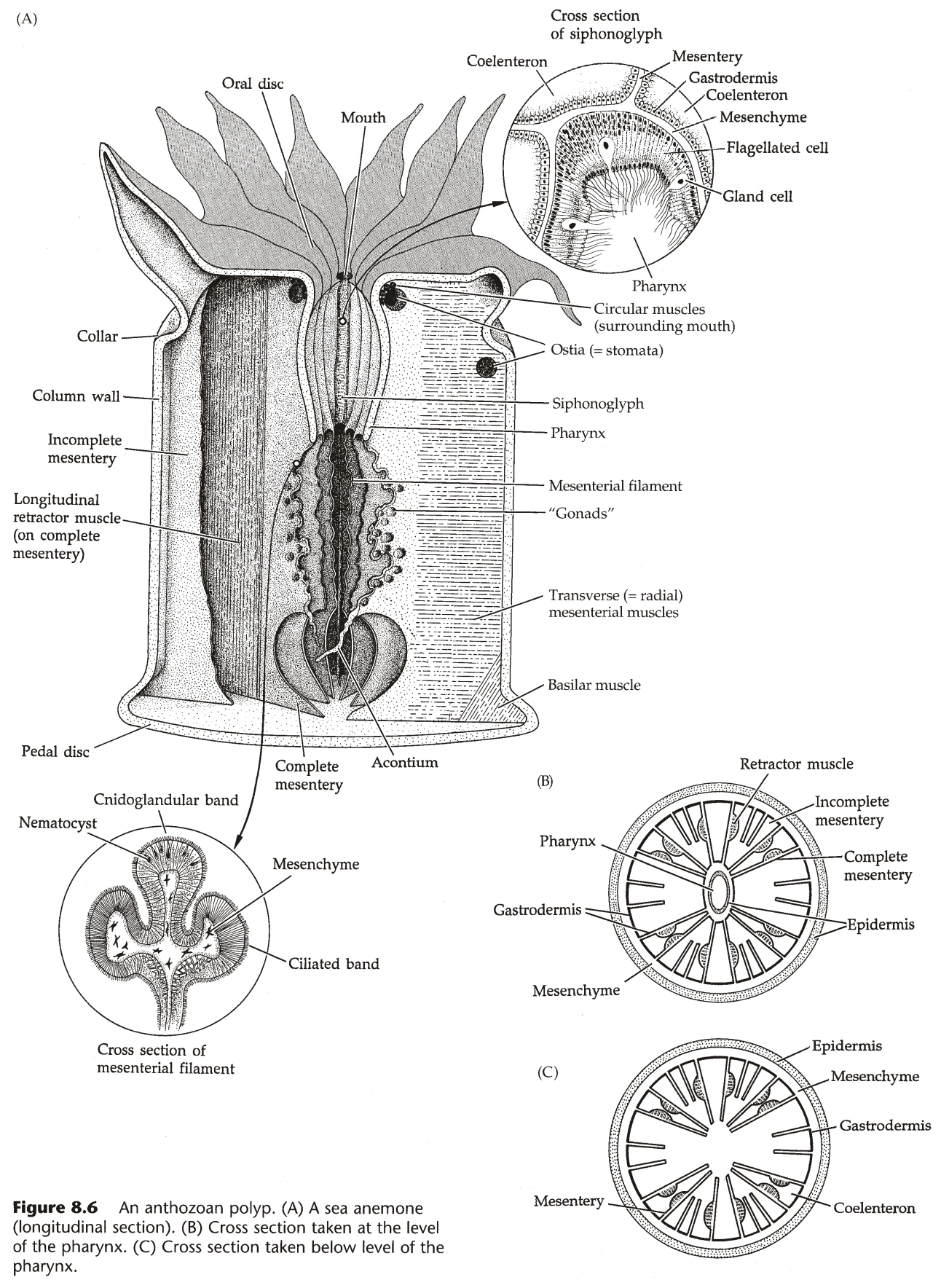
{kind=link}
图4:珊瑚纲的水螅型
{kind=link}
图5:螅茎的分枝方式

{kind=link}
图6:水母型
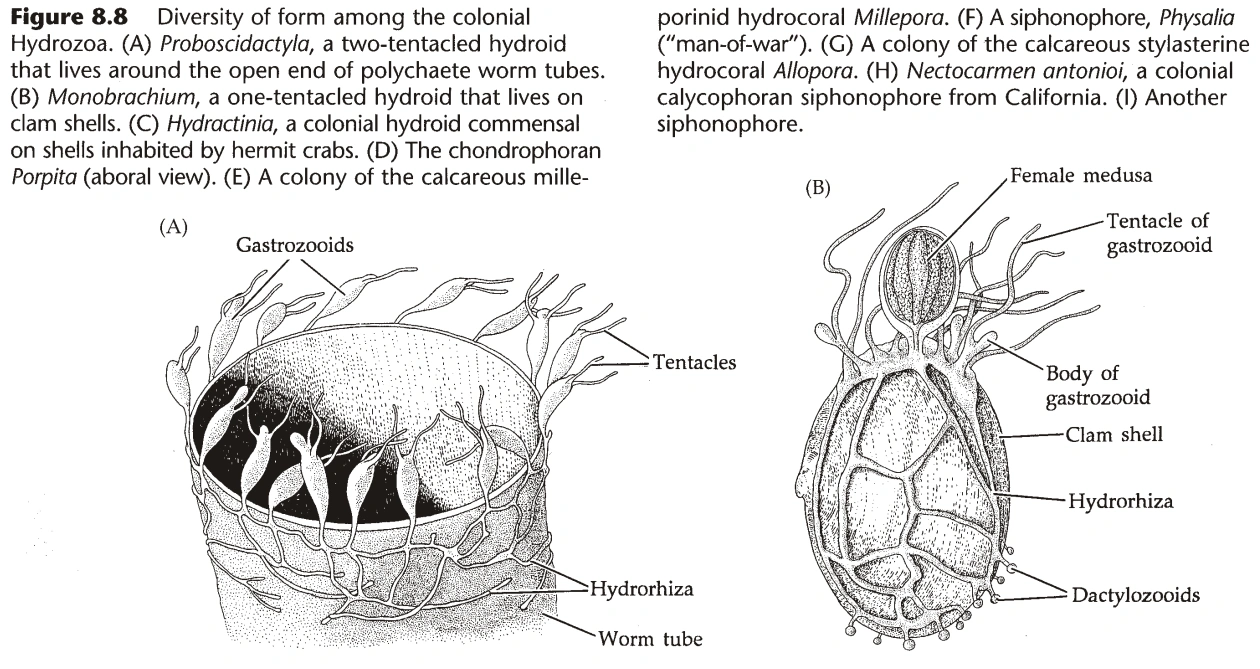
{kind=link}
图7A:水螅群体的多样形态
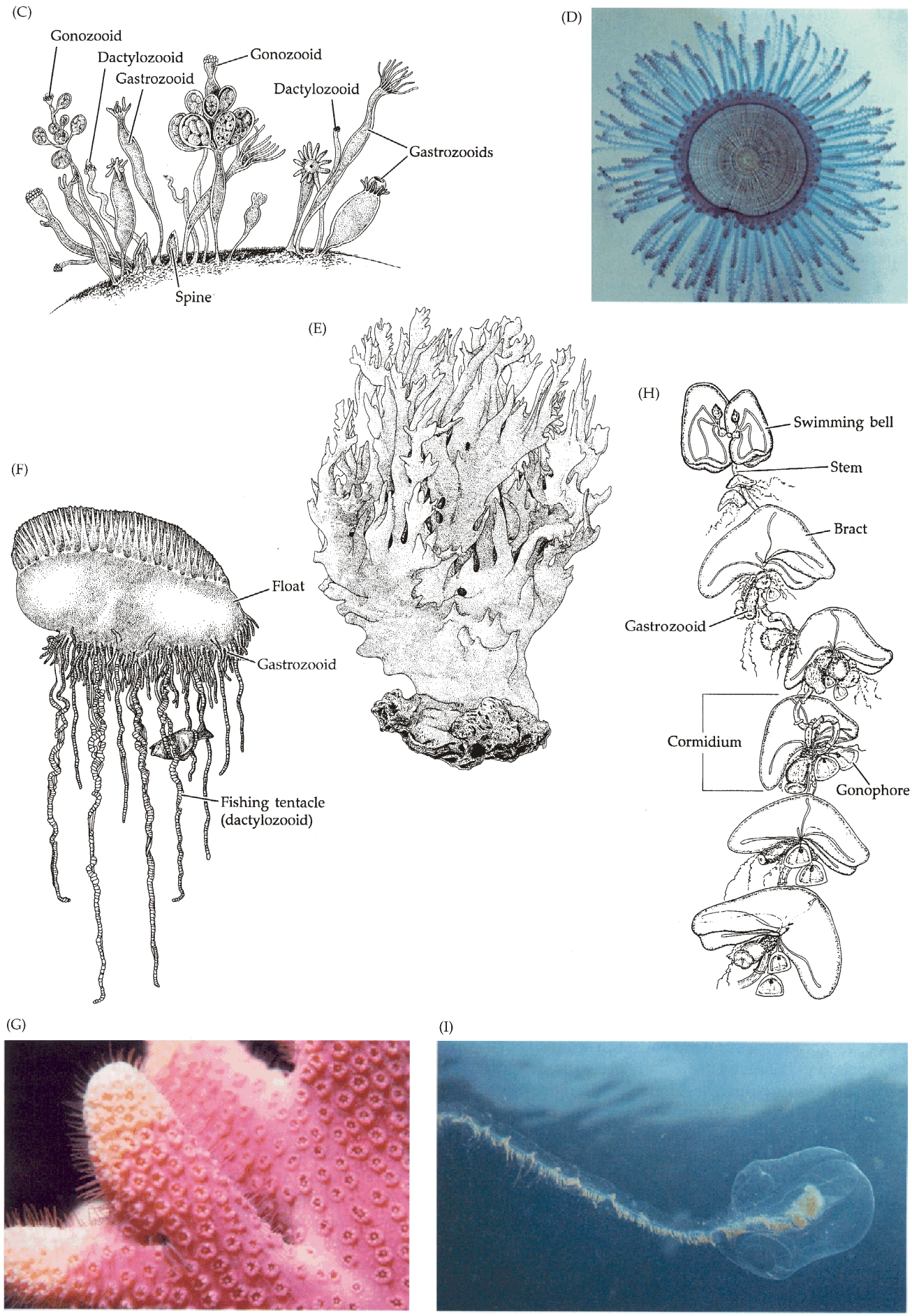
{kind=link}
图7B:水螅群体的多样形态


{kind=link}
图7C:珊瑚群体


{kind=link}
图7D:柳珊瑚群体
{kind=link}
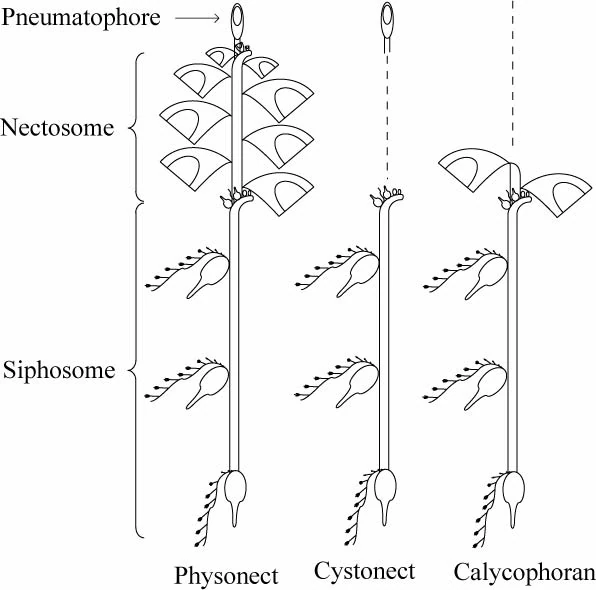
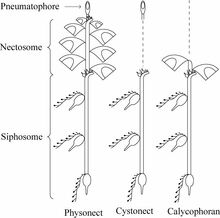
图8:管水母群体的三种形态

{kind=link}
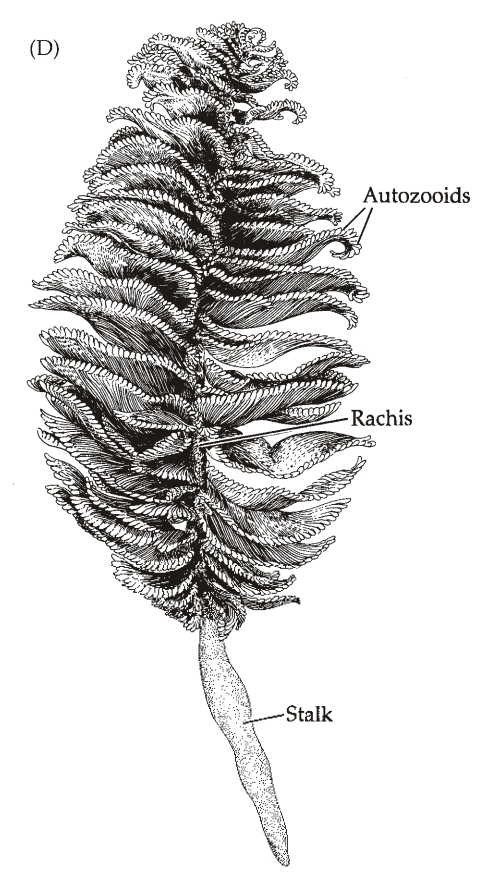
图9:海鳃的群体

{kind=link}
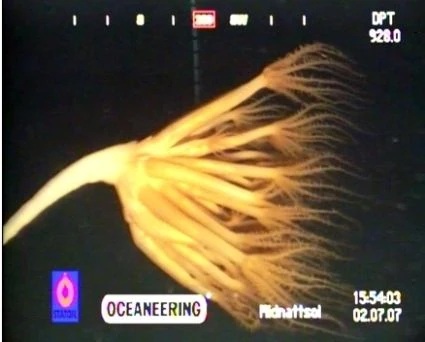
图10:伞形珊瑚
- 刺胞动物出现了最原始的肌肉细胞,称为肌皮细胞,构成刺胞动物的基本组织。(图1)
- 外胚层的肌皮细胞称为皮肌细胞(Epitheliomuscular Cell),内胚层的肌皮细胞称为营养肌细胞(Nutritive-muscular Cell)。
- 肌皮细胞的基部伸出管状结构,可收缩,称为肌线(Myoneme)。(图2)
- 肌线固着在来自外胚层的间质或中胶层上。
- 肌线有两种:轴向(Longitudinal)和环向(Circular)。
- 间质中有时有特化的肌皮细胞,已近似于肌肉细胞。
- 外胚层还含有感觉细胞、刺胞(Cnidocyte,又名刺丝囊,Nematocyst)、腺细胞、间细胞(Interstitial Cell)。
- 内胚层还含有刺胞和腺细胞。(水螅纲的内胚层无刺胞)
- 只有珊瑚纲有间质,其它刺胞动物只有中胶层,水螅纲的中胶层最薄。
- 刺胞是刺胞动物特有的防御工具,集中在嘴和触手(见下文)附近,组成Nematocyst Battery。
- 腺细胞大部分在内胚层,用于向细胞外(腔肠)分泌消化酶,内胚层的腺细胞集中在嘴附近,外胚层的腺细胞集中在基盘附近。
- 间细胞相当于干细胞,能分化成其它细胞类型。
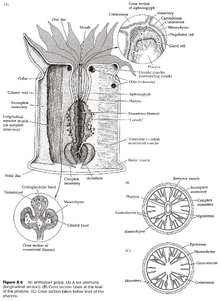
水螅型(图3、4)
- 所有刺胞动物都有水螅型形态,但在立方水母纲和钵水母纲中退化。
- 水螅型能发生无性生殖(出芽生殖),能形成群体,形态上比水母型多样。
- 反口部可为基盘(Basal Disc,基本形态)、足盘(Pedal Disc,部分珊瑚纲)、圆基(Physa,部分珊瑚纲)。
- 水螅纲的嘴在垂唇(Hypostome)上;珊瑚纲没有垂唇,但有口盘(Oral Disc)。
- 珊瑚纲的嘴有咽(Pharynx)与之相连,咽来自内胚层,咽壁上有小沟(Siphonoglyph),沟壁有纤毛,能将水推入腔肠。(咽属于外胚层)
- 嘴的边上常有触手(Tentacle),珊瑚纲的触手是腔肠的延伸,水螅纲的触手则是单层内胚层细胞。
- 触手的末端可逐渐变细(丝状触手,Filiform),也可膨大含大量刺胞(锤形触手,Capitate)。
- 触手可分枝,在八放珊瑚亚纲(Octocorallia)中每个水螅型个体有8个触手,每个触手有大量羽状分枝(羽枝,Pinnule)。
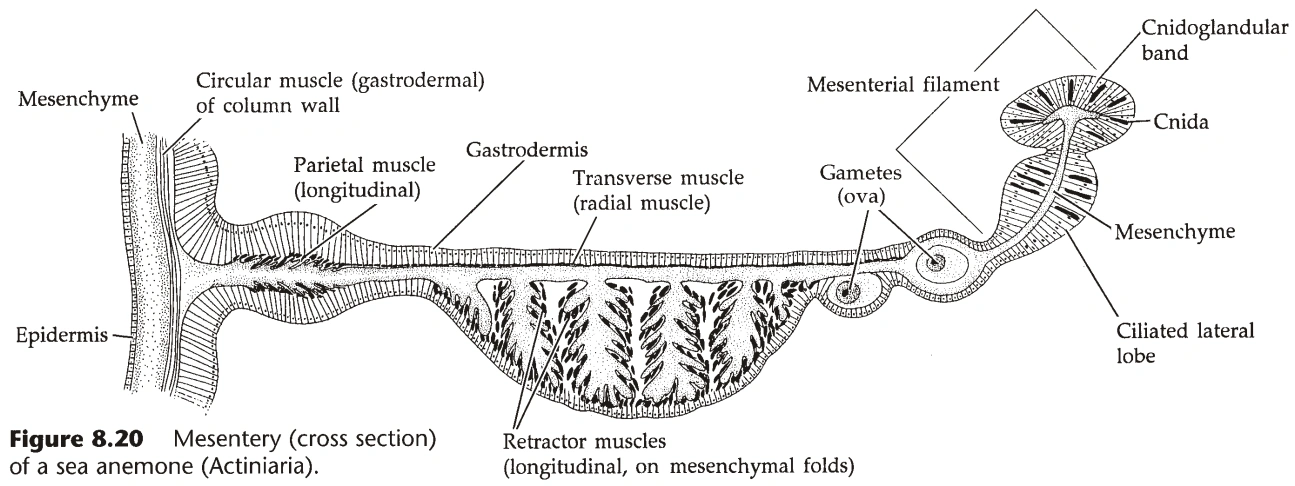
- 水螅纲的腔肠不分区,钵水母纲的腔肠被系膜(Mesentery)部分隔成四个腔,珊瑚纲的腔肠被系膜隔为很多个腔。(系膜的两侧都是内胚层,系膜的功能是增大消化面积)
- 珊瑚纲中,系膜未与咽相连的部分,其边缘增厚,有刺胞、腺细胞、纤毛,称为隔膜丝(Mesenterial Filament)。
- 部分海葵中,隔膜丝上能长出长线状物,称为枪丝(Acontium)。
- 珊瑚的不同分区间可通过管系(Solenium)相连。
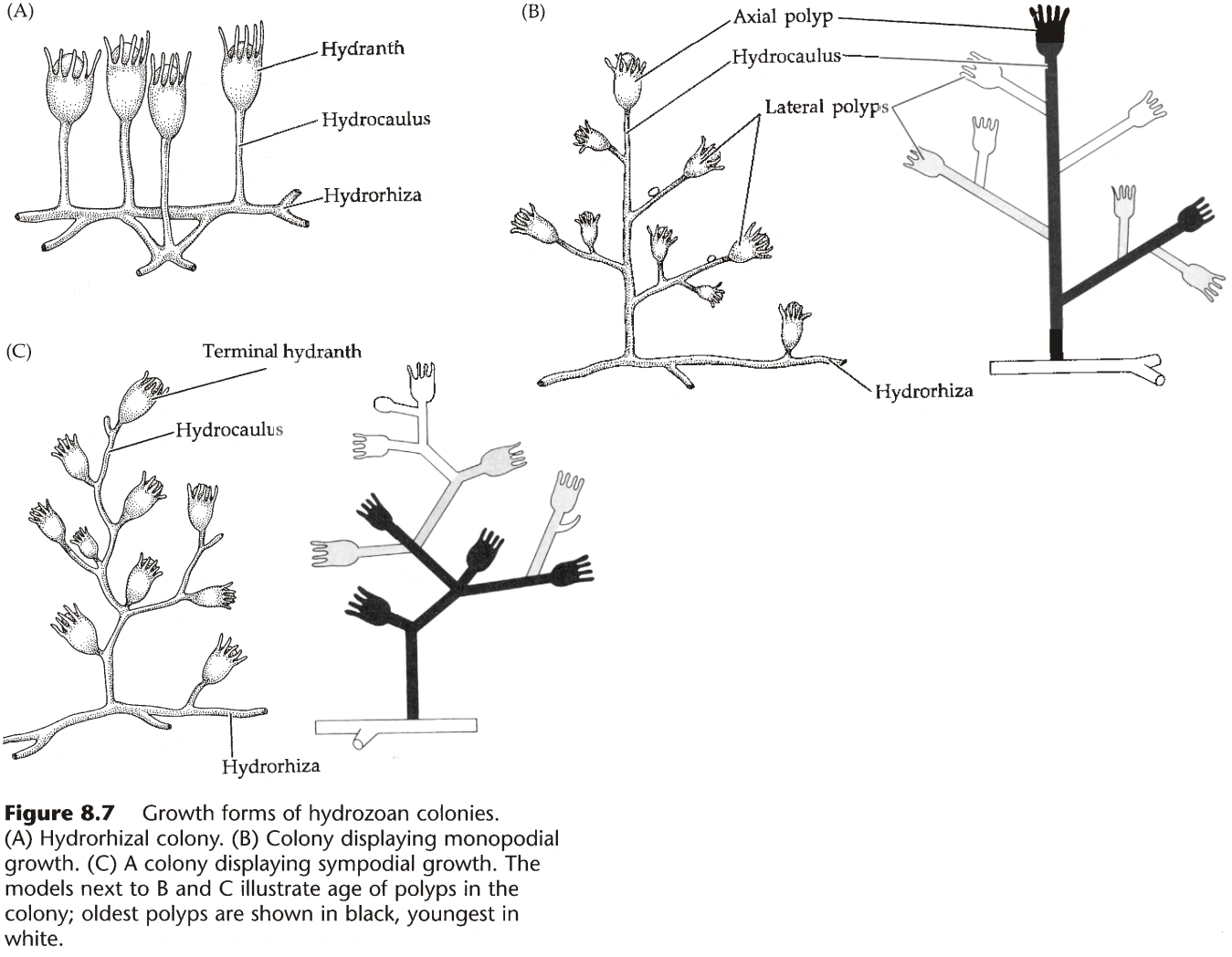
- 水螅纲的群体结构:
- 群体通过螅根(Hydrorhiza)固定在地面,从螅根上长出螅茎(Hydrocaulus)。
- 螅茎的末端,长出水螅的个体,称为水螅体(Hydranth),其腔肠与螅茎联通。
- 螅茎的活组织称为共肉(Coenosarc,又名Coenenchyme),外部有围鞘(Perisarc)包被,围鞘无细胞,由蛋白质和几丁质组成。(淡水中的水螅无围鞘)
- 部分水螅中,围鞘延伸至水螅体外,称为螅鞘(Hydrotheca)。
- 螅茎的分枝方式分单轴分枝(Monopodial)和合轴分枝(Sympodial)。(图5,参考维管植物的结构)
- 部分水螅体形态特殊:指状水螅体(Dactylozooid)含大量刺胞,专用于防御;生殖水螅体(Gonozooid)专用于繁殖。(普通的水螅体称为营养水螅体,Gastrozooid)。
- 指状水螅体常围着营养水螅体一圈,除防御外还有辅助捕捉食物的功能。
- 有螅鞘的生殖水螅体称为生殖壶(Gonangium)。
- 生殖水螅体的活组织称为子茎(Blastostyle)。
- 对于无世代交替的水螅,生殖水螅体产生孢子囊(Sporosac),孢子囊产生配子;对于有世代交替的,它产生水母型子代,子代既可脱离子茎,也可固着在子茎上,称为生殖体(Gonophore)。
- 管水母目(Siphonophora)有特殊的气胞囊(Pneumatophore,一种水螅型),含有气腺,能释放与空气成分相近的气体,使群体在水中上浮。
- 僧帽水母(Physalia)的气胞囊释放的气体含大量一氧化碳。
- 部分管水母除了气胞囊还有一种特殊的水母型结构,称为泳钟(Nectophore)。(图7、8)
- 钟泳亚目(Calycophorae)的泳钟长在螅茎的顶部,螅茎上长出若干合体节(Cormidium),每个合体节相当于一个小群体。
- 胞泳亚目(Physonectae)的螅茎顶部是一个气胞囊,后方跟有一串泳钟。
- 囊泳亚目(Cystonectae)的螅茎顶部只有气胞囊而无泳钟。
- 合体节可以从螅茎上脱离,在另一处固定生长,仍称合体节(Eudoxid)。
- 合体节内部的生殖体也可能发挥泳钟的功能。
- 一般珊瑚纲的群体结构类似水螅纲,但水螅型非常小,并且没有生殖体(原因见下文)。(图7)
- 柳珊瑚目(Gorgonacea)的分枝极多,既可呈灌木状也可只在一个平面内,形状明显受水流影响,即分枝方向几乎垂直于水流方向。(若局部水流有两种流向交替,则柳珊瑚可在两个平面内分枝)
- 海鳃目(Pennatulacea,属于珊瑚纲)的群体结构:(图9)
- 群体的基础是一个巨大的的水螅型,一部分埋在地下(Stalk),一部分露出地面(Rachis)。
- 露在地上的部分出芽形成大量水螅型。
- 一群水螅型在主干上呈行或呈螺旋排列,有时联合形成“叶”。
- 水螅型有两种:营养水螅体(Autozooid)同上;管状水螅体(Siphonozooid)较小,负责创造水流。
- 伞形珊瑚(Umbellula)的水螅型全部长在基础水螅的顶端,呈伞状。(图10)
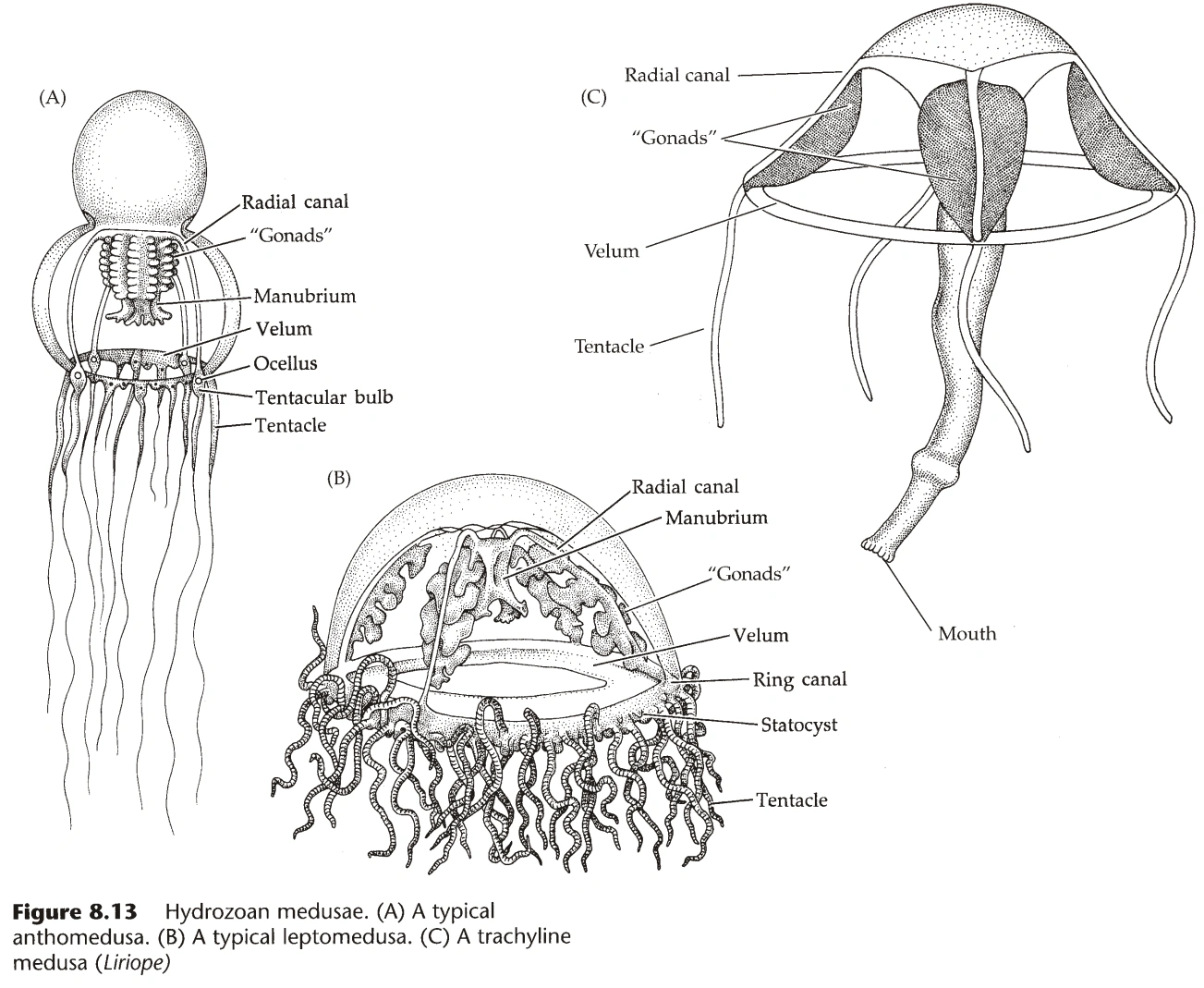
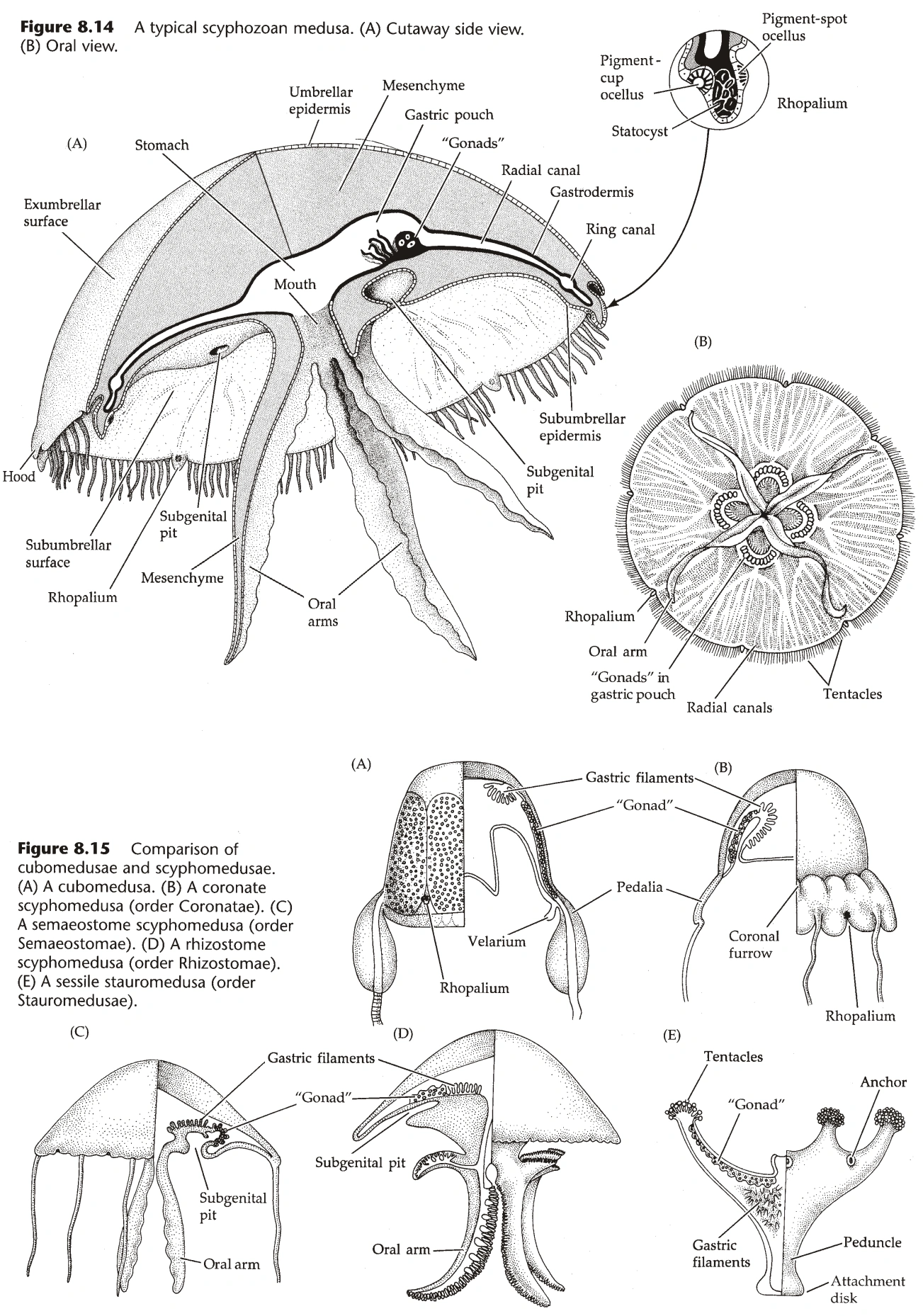
水母型(图6)
- 珊瑚纲无水母型,其它三个纲都可有水母型。(故珊瑚纲无生殖体)
- 水母不能组成群体,形态没有水螅型多样。(但能以生殖体的形式参与群体,见上文)
- 绝大部分水母能自由运动,极少数固着海底。
- 水母型的组织结构与水螅型相同。
- 水母有一个伞状的顶,顶的上侧称为外伞(Exumbrella),下侧称为内伞(Subumbrella,又名下伞)。
- 水母的嘴开在内伞的中央,通常此处延伸出一个柱状结构,称为口柄(Manubrium,又名垂管)。
- 腔肠在伞的内部,伞通常向外辐射状伸出若干辐管(Radial Canal),是腔肠的延伸。
- 辐管在底部通过环管(Ring Canal)汇合,环管有时向内延伸出扁平结构,称为罩膜(Velum)。
- 水母的触手长在环管下。
- 辐管数量通常是四的倍数,因此水母型一般是四射对称(Quadriradial Symmetry)。
生理学
{kind=link}
图11:珊瑚(实际是珊瑚纲动物分泌的钙质骨骼)

{kind=link}
图12:刺胞

{kind=link}
图13:系膜上的肌皮细胞

{kind=link}
图14:刺胞动物的运动

{kind=link}
图15:刺胞射出
{kind=link}
图16:水母有时也能出芽生殖
{kind=link}
图17:钵水母和立方水母,注意生殖腺的位置不同


{kind=link}
图18:节片生殖
支持
- 水螅型和水母型都主要通过间质或中胶层支持身体,水母较为柔软。
- 水螅型主要通过水压将自己固定在地面上。
- 珊瑚的间质中有纤维,可增强体壁强度。
- 部分珊瑚能将固着的地面的物质融入围鞘,增强体壁。
- 一些水螅型群体的共肉中有坚硬的骨骼结构。
- 骨骼结构有三种:轴向骨骼(Axial Skeleton)、钙质骨针(Calcareous Sclerite)、钙质网络(Calcareous Network)。
- 轴向骨骼是共肉内壁的变形细胞(Amebocyte)向共肉的腔分泌的,主要成分为蛋白质和粘多糖。(糖类具体成分不明)
- 轴向骨骼可以很坚硬,如黑珊瑚(Antipatharia)的轴向骨骼可以制成珠宝。
- 钙质骨针是间质中的成骨细胞(Scleroblast)分泌的碳酸钙盐,颜色和形态多样。
- 如果钙质骨针相连形成一个整体,就形成了钙质网络。
- 最常见的钙质网络就是珊瑚纲动物分泌的珊瑚(Coral)。
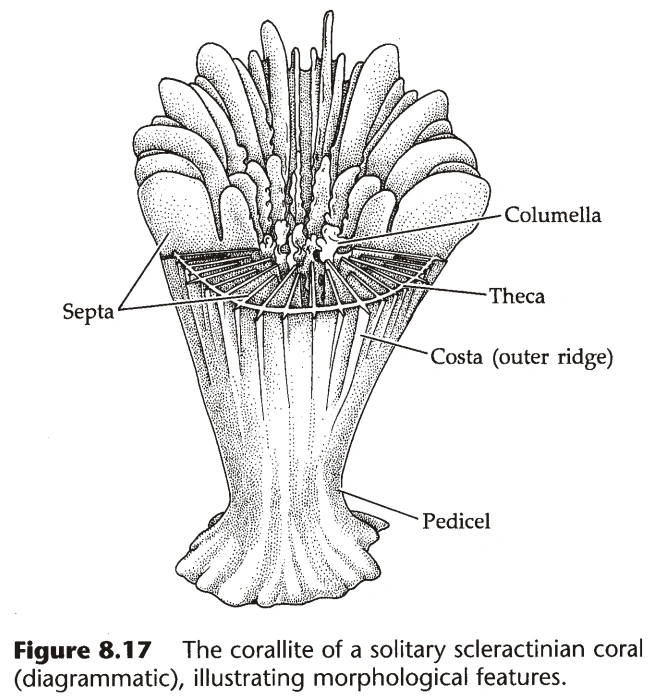
- 整个群体分泌的钙质网络称为珊瑚体(Corallum),属于每个个体的部分称为珊瑚单体(Corallite)。
- 珊瑚单体中,外壁称为Theca,系膜的骨骼称为Septa。
营养
- 大部分刺胞动物是肉食性的。
- 刺胞(触手、指状水螅体)将猎物杀死或麻醉,再被水流送入腔肠,触手可运动将食物推入腔肠。
- 一些来自食物的物质可刺激触手运动和嘴张开,如谷胱甘肽和自由氨基酸。
- 腔肠内,腺细胞分泌的消化酶将猎物初步分解,多肽、脂质、糖类通过胞吞和胞饮进入营养肌细胞。
- 消化在食物泡中完成,废物通过胞吐进入腔肠,通过嘴离开。
- 部分珊瑚有枪丝,含大量刺细胞和腺细胞,辅助消化。(若珊瑚受到猛烈碰撞,枪丝可被射出嘴部,起防御功能)
- 部分珊瑚能在表面分泌黏液,粘住食物,再通过纤毛送入嘴中。(黏液本身可作为鱼类等的食物)
- 氧气直接从水中获取,埋在地下的部分发生无氧呼吸。
- 含氮废物以氨的形式排放。
防御
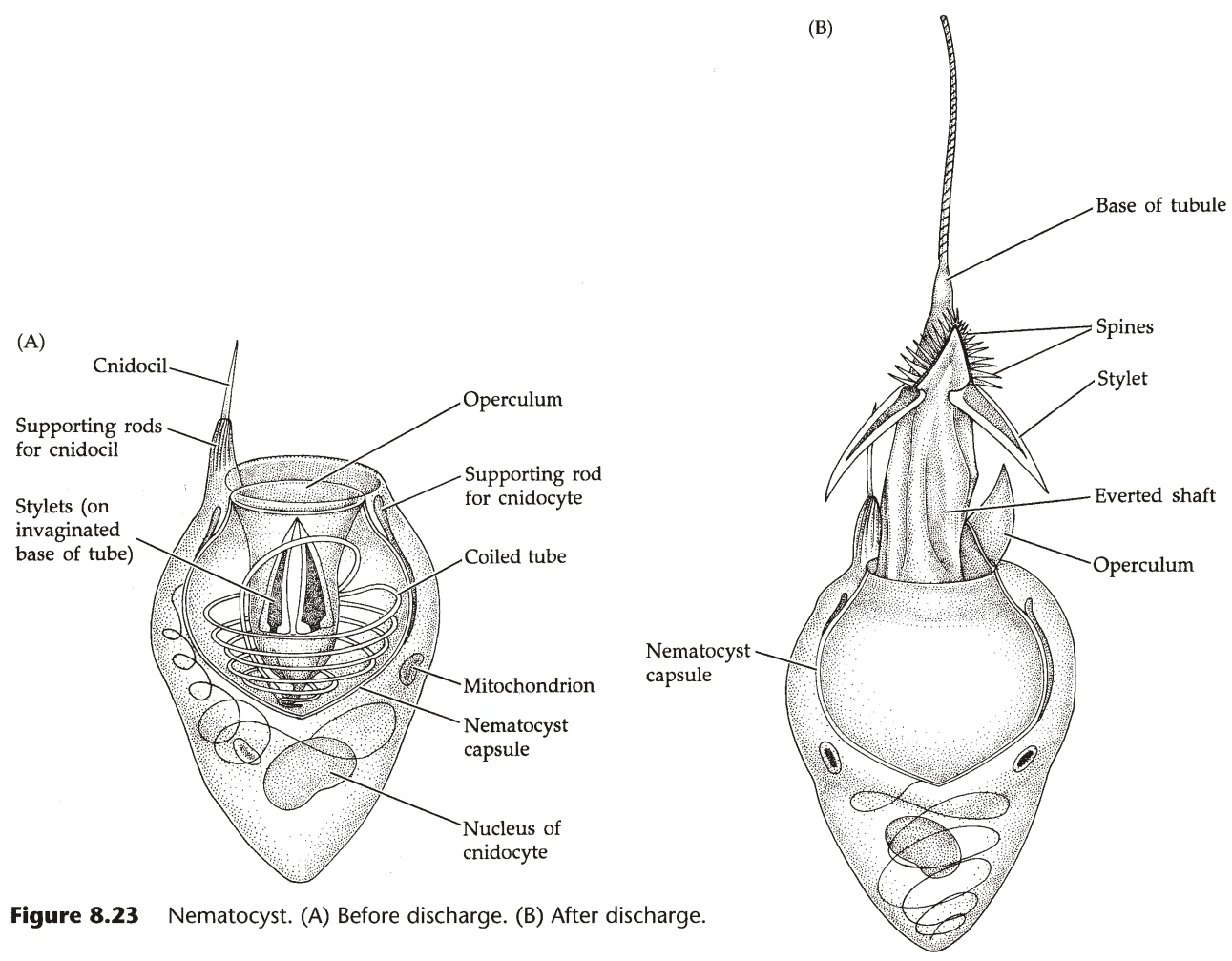
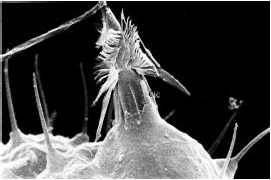
- 刺胞动物的防御主要是通过特有的刺胞完成的。
- 刺胞中的特殊结构是刺丝泡(Cnidae)。
- 刺丝泡有一层荚膜,成分是类似胶原蛋白的蛋白质。
- 荚膜的一头向内凹陷,从凹陷的尖端长出刺丝,围绕凹陷的荚膜螺旋。(凹陷荚膜长有口锥(Stylet)结构,功能不明。)
- 凹陷口处常有盖结构(Operculum),避免刺丝不必要时弹出。
- 刺死的结构尚未搞清楚,可能是高尔基体分泌的,也可能是远古细菌内共生带来的。
- 刺丝的末端是针,针尖有小孔,可注射毒物(蛋白质、酚类物质)。(有些物种能释放海葵毒素(Palytoxin),是已知最强的毒物之一,它可能是与之共生的细菌合成的)
- 刺胞表面有一个刺针(Cnidocil),是机械感受器。
- 荚膜中含大量钙离子,受到刺激时,钙离子被大量释放入细胞质,导致渗透压上升,水大量涌入细胞,将凹陷的荚膜推出细胞。
- 刺胞弹出时,最大加速度可达五百万g。
- 特殊的刺胞:
- 六放珊瑚目(Zoantharia,属于珊瑚纲)含有旋胞(Spirocyst),其刺丝没有针,用于粘住猎物。
- 角海葵目(Ceriantharia,属于珊瑚纲)含有黏胞(Ptychocyst),其刺丝没有针,不是用于捕捉食物,而是用于编织成一个管状结构,作为保护组织。
运动
- 刺胞动物的运动主要是通过肌皮细胞完成的。
- 珊瑚纲的肌皮细胞最为发达(尤其是海葵),间质有大量特化的肌皮细胞,已无皮肤功能。
- 海葵的大部分肌皮细胞来自内胚层,外胚层的肌皮细胞集中于口盘和触手处。
- 肌皮细胞的肌线分轴向和环向,轴向收缩使个体长度变短,环向收缩使腔肠直径变窄。
- 珊瑚的系膜上还有径向肌线,收缩使系膜变短,促进食物进入腔肠。(图13)
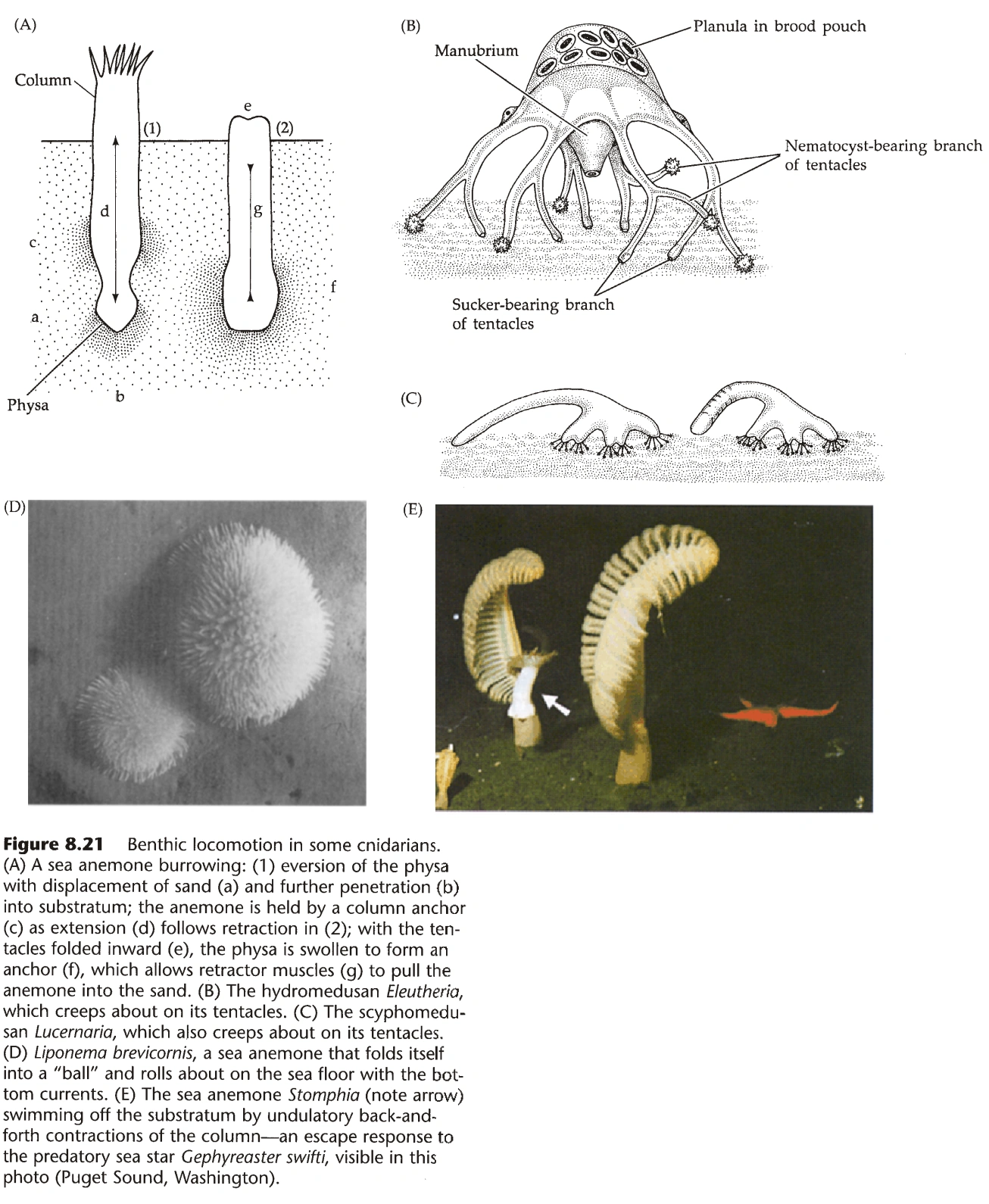
- 大部分水螅型是固着的,不能运动,有些不成群体的水螅型能以翻跟斗的方式运动。
- 很多水母型能以触手为“脚”,在地面爬行。(图14)
生殖
- 刺胞动物再生能力很强,部分物种中取下外胚层少数细胞即可长成一个新个体。
- 曾有人将一个水母型翻过来,即内胚层翻至外面,结果个体仍能存活和生殖,内外胚层功能自动交换。
- 水螅纲:
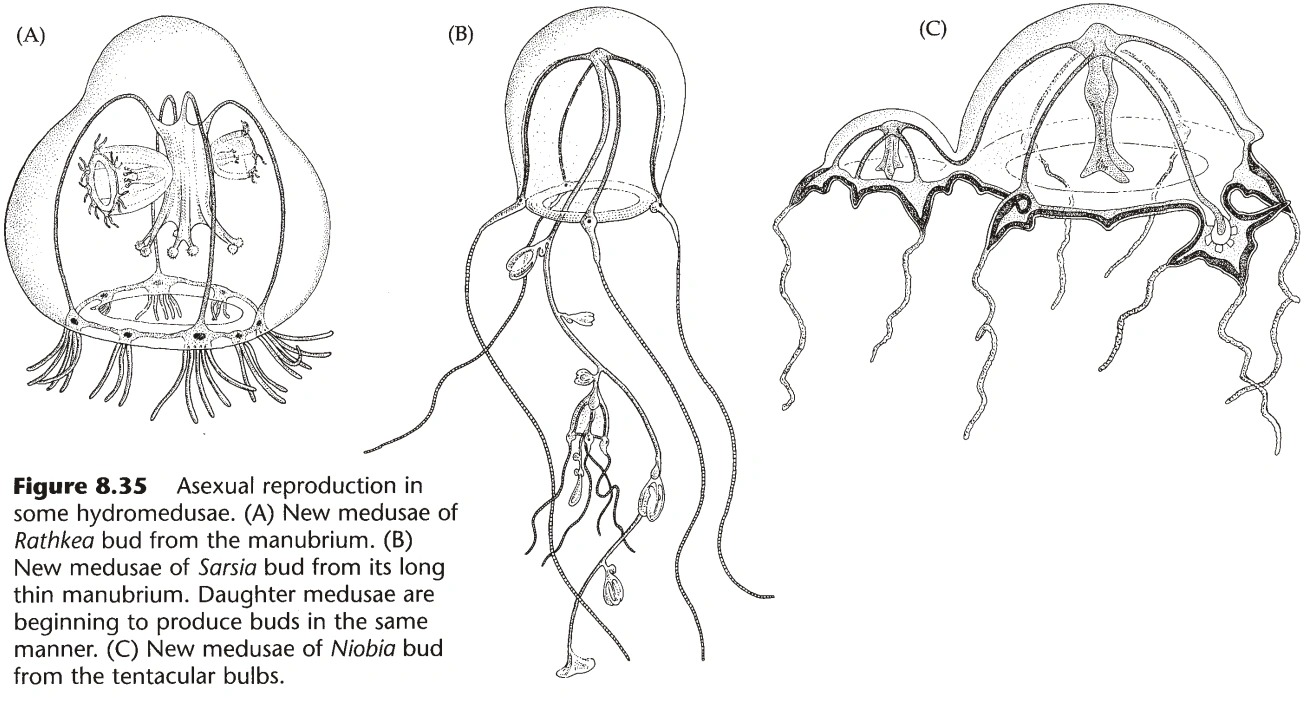
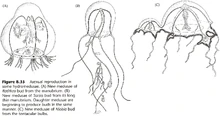
- 水螅型出芽生殖产生水螅型或水母型,水母型有时也能出芽生殖产生水母型。(图16)
- 水螅无水母型,产生孢子囊(源于外胚层);而数枝螅可以产生水母型。
- 水母型中,大部分雌雄异体,配子细胞源于外胚层,性腺在口柄上、辐管上或下伞上。
- 珊瑚纲:
- 无水母型,水螅型无性生殖方式多样,有轴向二分裂(Longitudinal Division)、足部撕裂(Pedal Laceration)、横向二分裂(Transverse Division)、触手出芽生殖、单性生殖(产生浮浪幼虫)。
- 有性生殖也存在,既可雌雄同体也可雌雄异体,有些群体可杂性同株。
- 配子细胞源于系膜上的内胚层。
- 钵水母纲和立方水母纲:
- 水螅型已退化,很小,但仍能形成群体,仍能出芽生殖。
- 钵水母的水螅型通过节片生殖(Strobilation)产生多个水母型,立方水母的水螅型则直接变态成为水母型。
- 节片生殖直接产生的未成熟的水母型称为蝶形幼体(Ephyra)。
- 水母型中,性腺来自内胚层,在腔肠内,钵水母的性腺和消化腺很近,立方水母中二者分离。
附录:栉水母动物门
{kind=link}
图19:球栉水母
{kind=link}
图20:栉水母口道分支解析
{kind=link}
图21A:其它栉水母的形态学
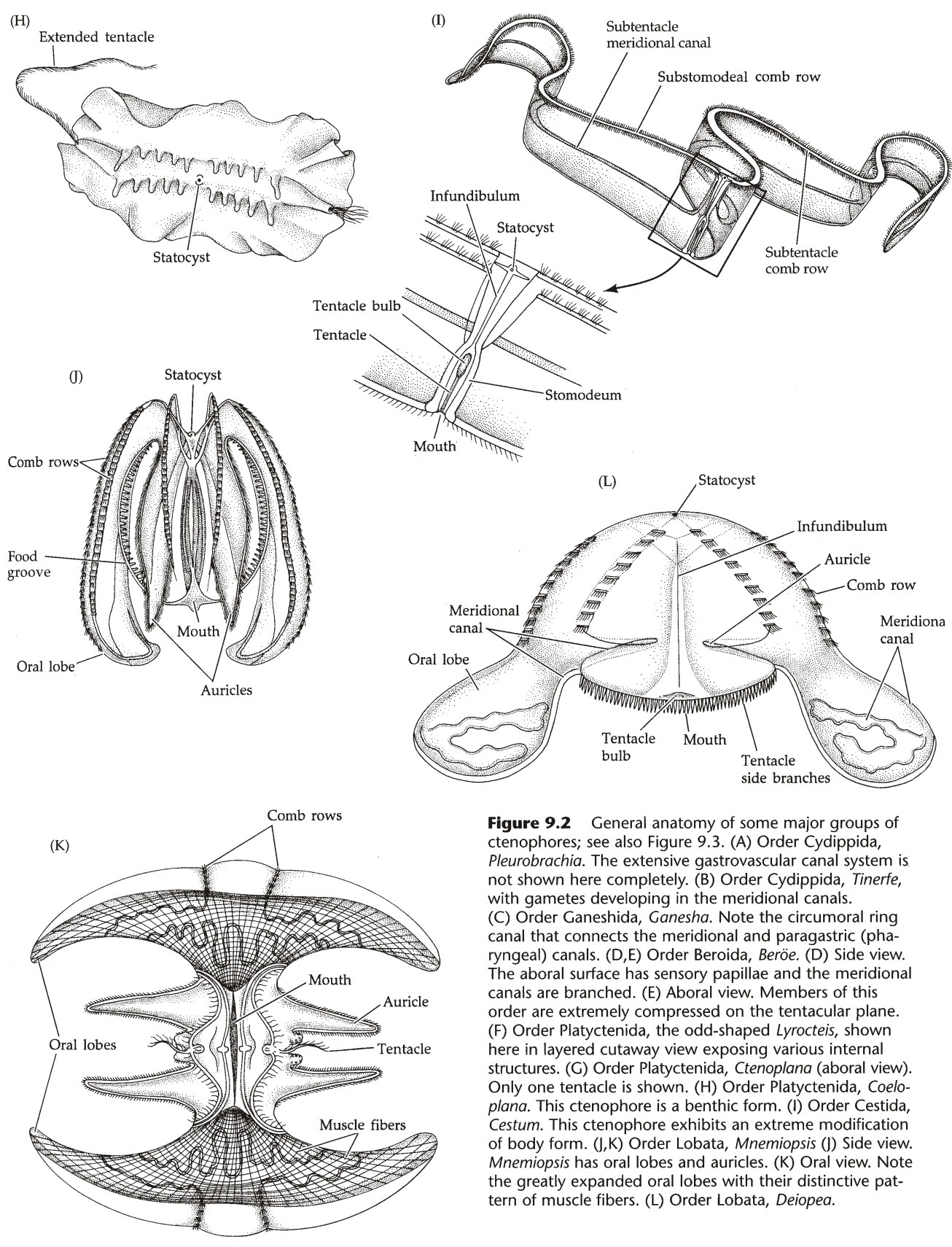
{kind=link}
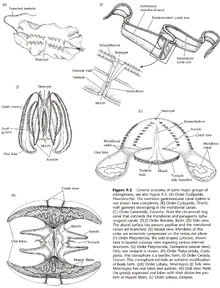
图21B:其它栉水母的形态学

{kind=link}
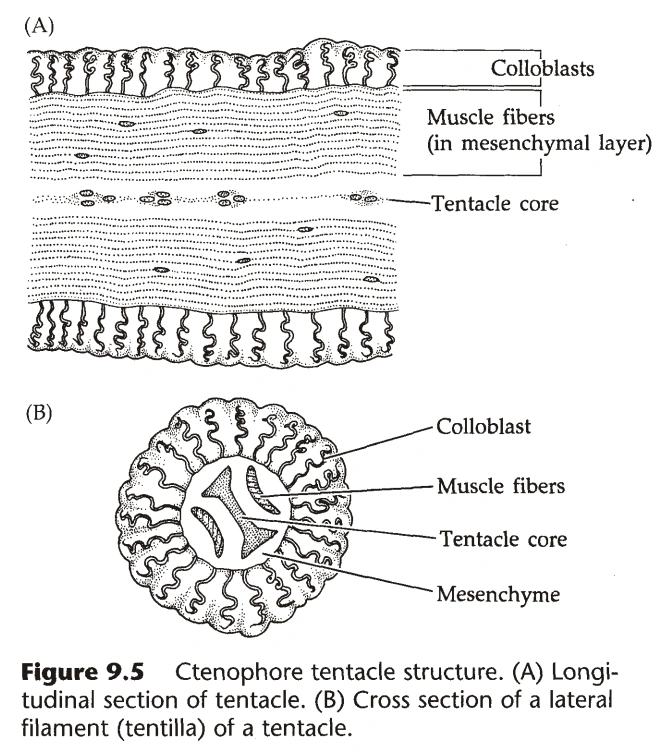
图22:触手的结构

{kind=link}
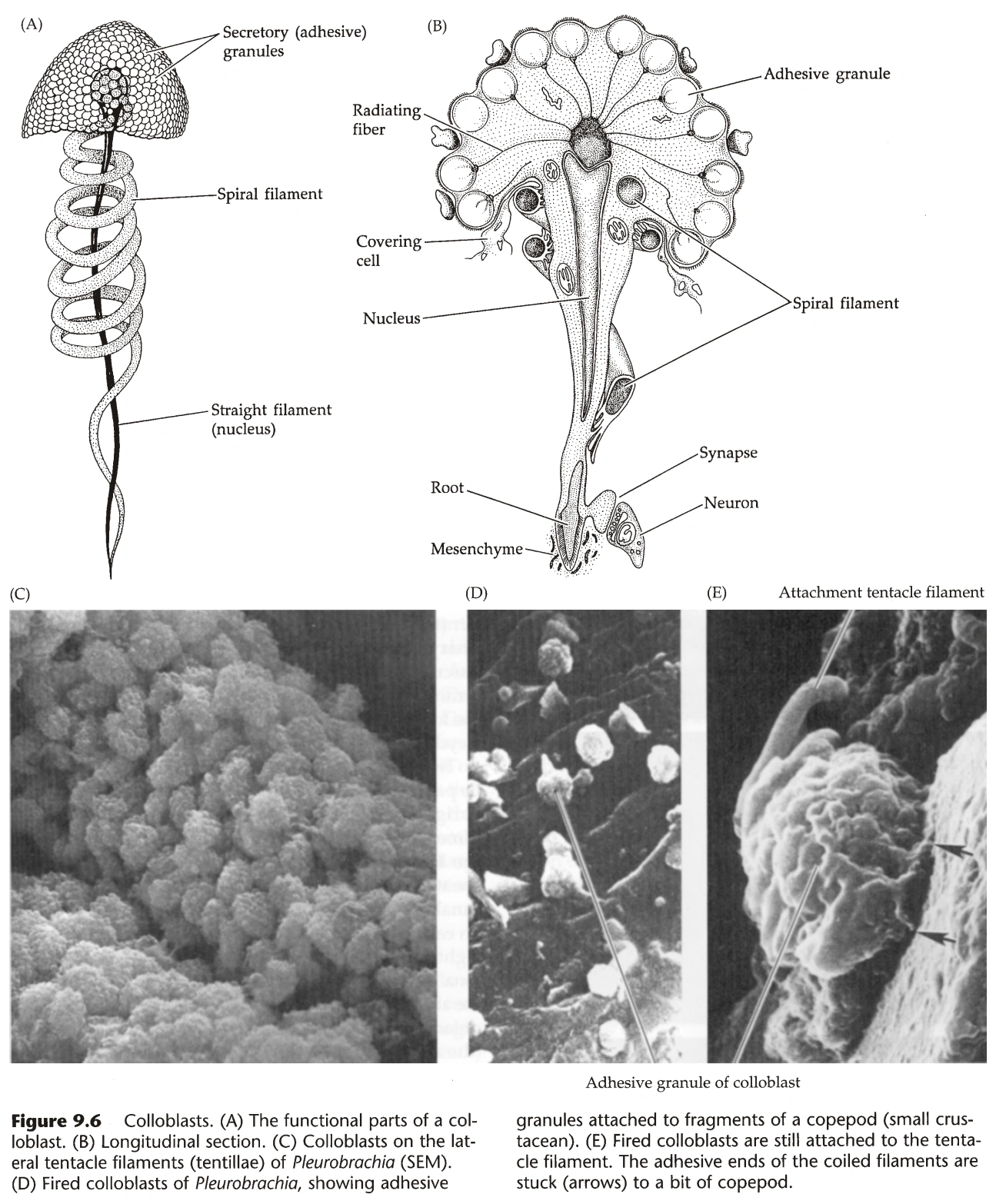
图23:粘细胞
- 栉水母动物与刺胞动物很相似,又是很小的一个门,故在此附录。
- 栉水母动物有没有三胚层仍具争议,但它确实都有间质。
- 栉水母动物没有miRNA,也没有Hox基因。
- 栉水母的内外胚层都是两层细胞。
- 栉水母没有世代交替。
- 栉水母的神经系统仍为原始的网状系统,但它的神经细胞被认为是独立进化出的,且有向栉板(见下文)靠拢的倾向。
- 目前认为栉水母是刺胞动物与高等动物分离后才分离出的
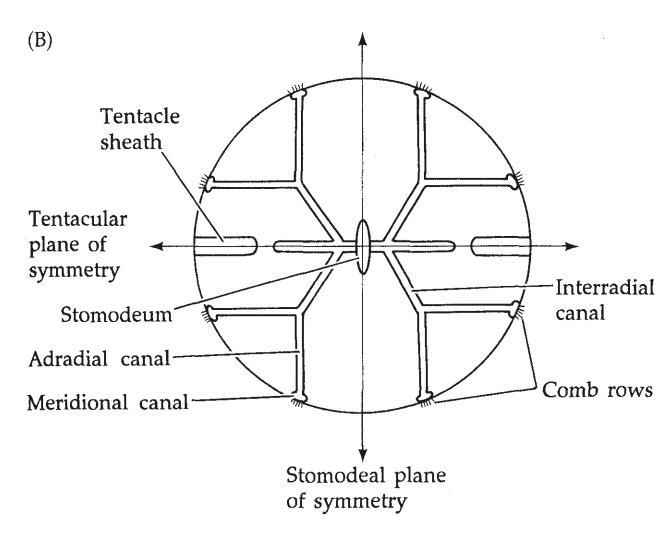
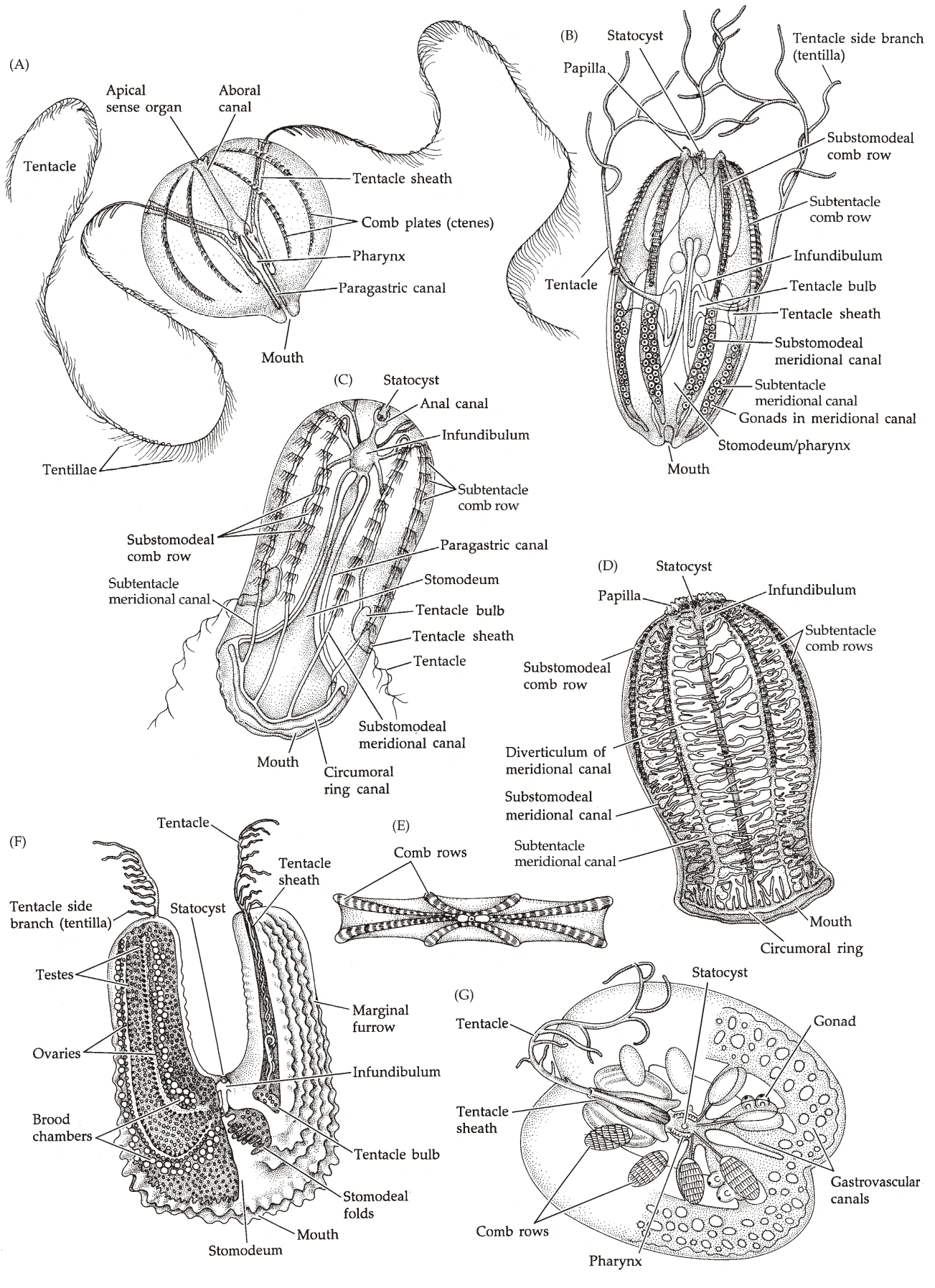
形态学(图19)
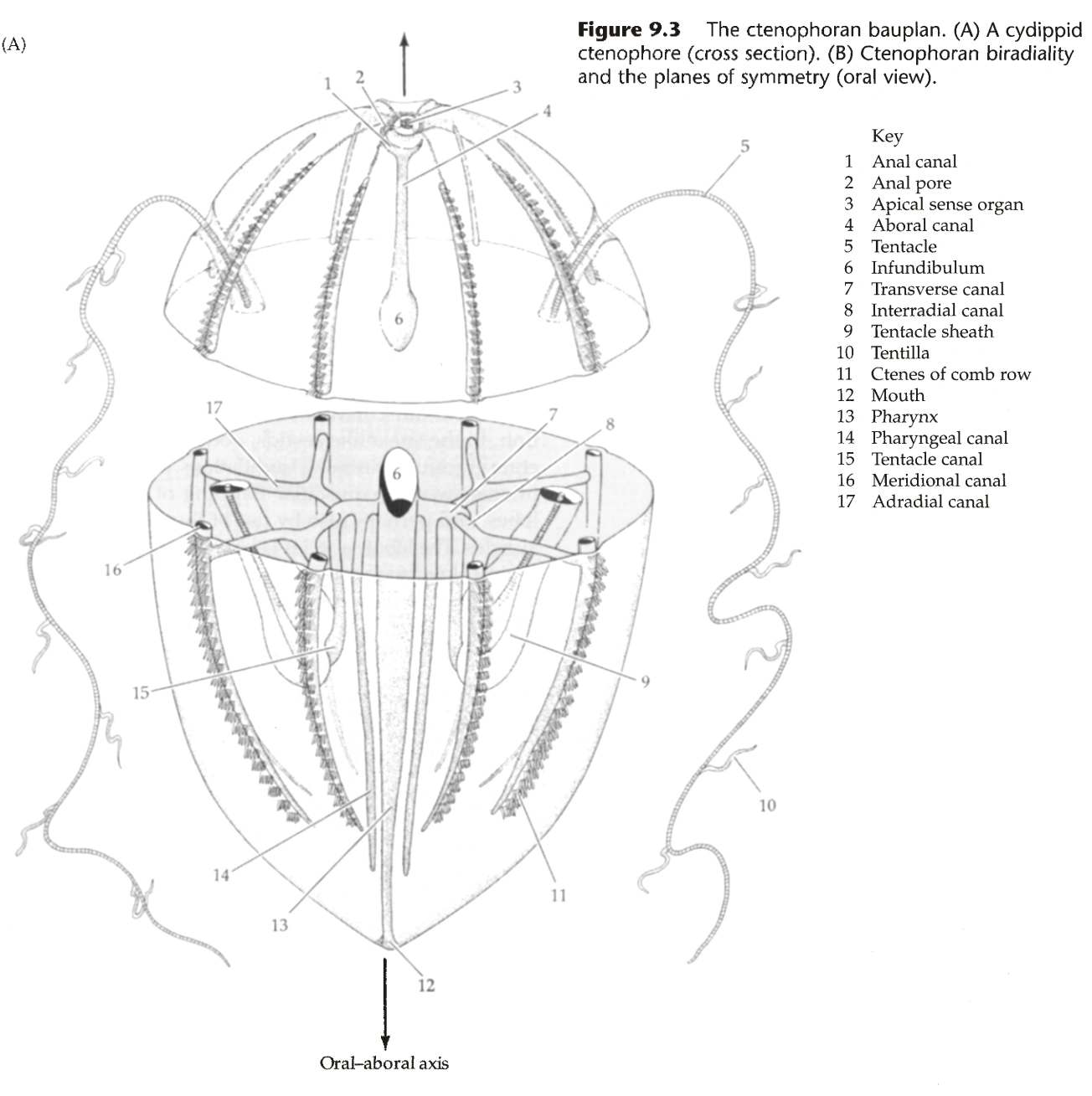
- 栉水母呈椭球形,体表有8条均匀分布的栉板(Comb Plate)。
- 栉板从口极伸向反口极,由大量纤毛组成。
- 很多物种中,身体两侧各有一个深的凹陷,称为触手鞘(Tentacle Sheath),外壁有纤毛,从中伸出触手。(因为有触手,栉水母动物一般是两侧辐射对称)
- 触手可伸出短分支,称为Tentilla;触手可通过触手鞘中的肌肉收缩。
- 触手没有刺细胞(极个别物种除外),但有大量粘细胞(Colloblast),有刺细胞则无粘细胞。
- 嘴的里面是口道(Stomodeum),在口道面(Stomodeal Plane)处变宽,之后直通反口极的肛门。
- 口道在口道面下方分支,先伸出横突管(Transverse Canal)。
- 横突管向下伸出咽管(Pharyngeal Canal)和触手管(Tentacle Canal),向栉板伸出间辐管(Interradial Canal),沿着栉板形成子午管(Meridional Canal)。
生理学
- 很多栉水母生活在深海,间质中的水分占身体绝大部分质量(可达96%)。
- 栉水母的肌肉细胞已大部分移动至间质中,虽然上皮细胞和肌肉细胞仍未分开。
- 栉水母都是肉食动物。
- 触手与浮游生物接触时,粘细胞弹出,释放大量黏性物质粘住猎物,然后通过肌肉运动送入口中。(相比于刺细胞的优点是不会破坏猎物)
- 粘细胞结构复杂,如图23,由分泌小泡、螺丝、直丝(特化的细胞核)组成。
- 也有些物种的粘细胞不会弹出,猎物被粘在体表后通过纤毛的运动送入口中。
- 有些栉水母以刺胞动物的触手为食,且能保留触手中的刺细胞,将它并入自己的外胚层用以防御,称为盗刺细胞(Kleptocnida)。
- 消化、呼吸方式类似刺胞动物。
- 栉水母的顶部有一个特殊的细胞,称为平衡石(Statolith),从边上伸出4束纤毛,称为平衡器(Balancer),从每束中伸出2根与栉板相连,称为纤毛沟(Ciliated Furrow或Ciliated Groove)。
- 栉水母向一侧倾斜时,平衡石对一侧栉板的压力增大,使此处栉板的纤毛加快运动,将个体调整回直立状态。
- 栉水母的再生能力仍很强。
- 大部分栉水母是雌雄同体,少数雌雄异体。
- 栉水母的性腺在子午管内壁上,配子细胞直接释放入海水,可自交。
- 卵子的周围有一群营养细胞(Nurse Cell)陪伴。
- 幼虫称为球栉水母幼虫(Cydippid Larva)。