总论
- 常见的细胞死亡方式有三种:细胞凋亡(Apoptosis)、细胞坏死(Necrosis)、细胞程序性坏死(Necroptosis)。
- 细胞凋亡是多细胞生物的一种正常的生理过程,目的在于控制发育,移除多余细胞。(主要出现于动物,植物中也有)
- 细胞坏死和细胞程序性坏死都是病理状态下的细胞死亡方式,都会引起炎症。
- 细胞自噬是一种和衰老联系在一起的细胞死亡。
细胞凋亡

{kind=link}
图1:胱天蛋白酶分启动者和执行者
{kind=link}
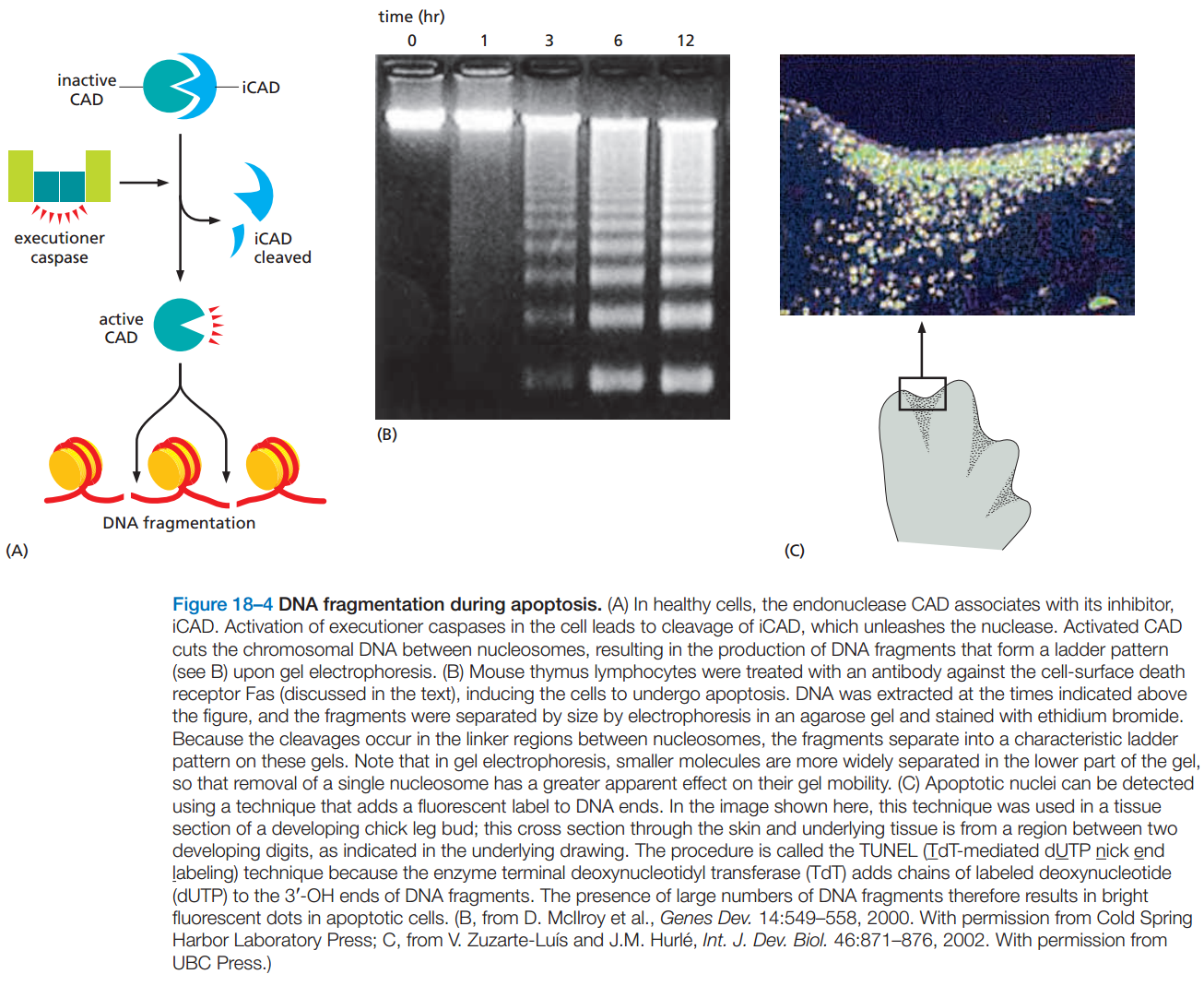
图2:细胞凋亡后的DNA呈梯状条带

{kind=link}
图3:细胞凋亡相关信号通路

{kind=link}
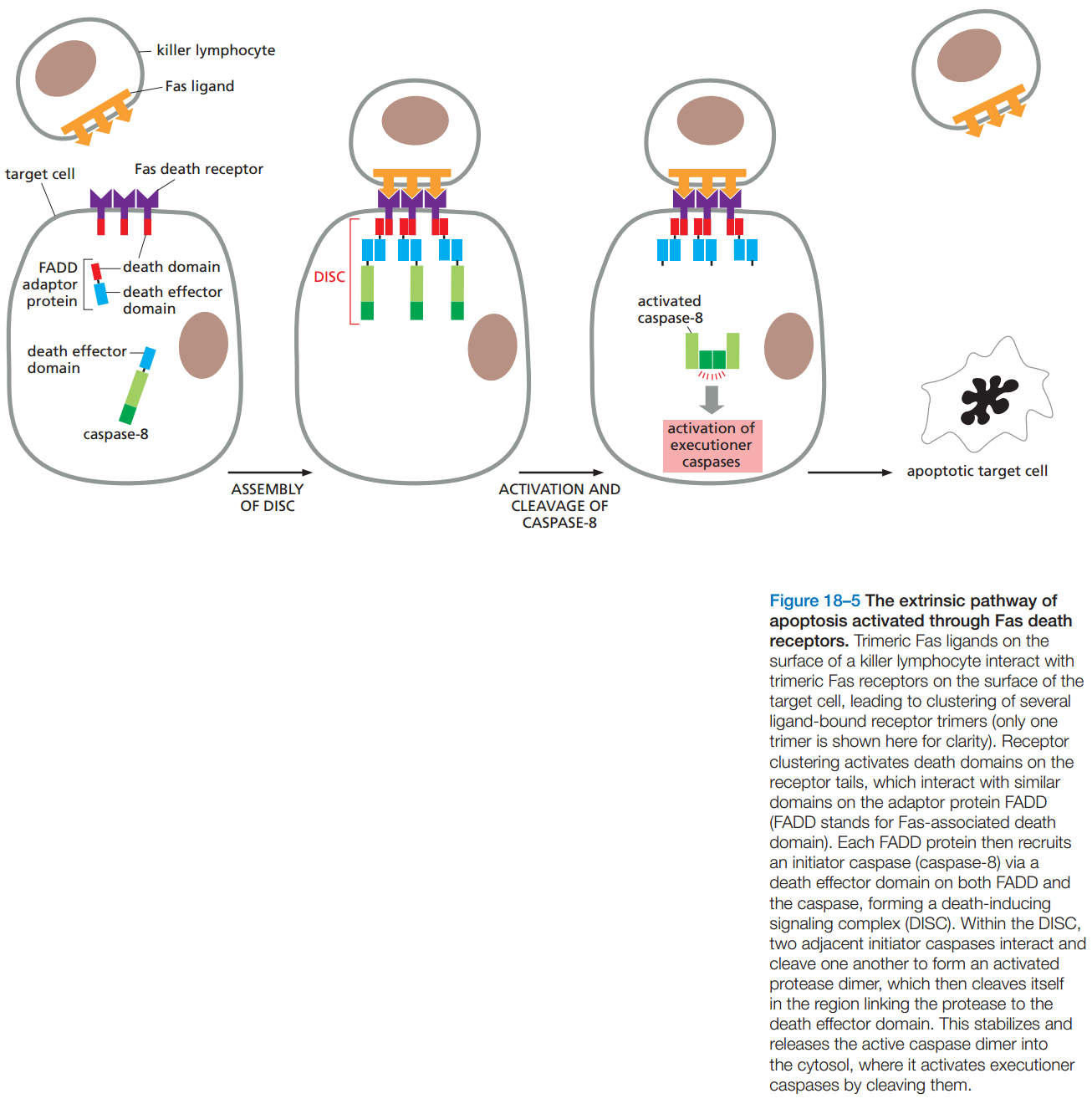
图4:外源途径


{kind=link}
图5:内源途径


{kind=link}
图6:生存因子的三种机制
- 细胞凋亡时的形态变化:
- 细胞收缩,细胞骨架坍塌,核膜解体,染色质浓缩并破碎。
- 细胞表面向外突出,然后破碎成很多凋亡小体(Apoptotic Body)。
- 细胞周围的吞噬细胞迅速将凋亡小体消化,细胞内容物不会溢出,不发炎。
- DNA的降解是通过CAD核酸内切酶完成的,它只在核小体之间切割,所以切出的DNA用凝胶电泳法按长度分离时,会呈现明显的梯状条带。(图2)
- 磷脂酰丝氨酸(Phosphatidylserine)一般只存在于细胞膜内叶,凋亡时进入外叶,这会吸引吞噬细胞。
- 细胞凋亡的关键是一类蛋白酶,称为胱天蛋白酶(Caspase),特征是活性中心有半胱氨酸,且切割位点含天冬氨酸,进化上它们高度保守。(参见常见酶促反应类型)
- 一般时期,胱天蛋白酶以无活性的前体存在;凋亡时,它们通过级联反应被激活。
- 胱天蛋白酶分两类:启动者(Initiator)和执行者(Executioner);启动者在信号通路中被激活,并激活执行者,由执行者降解细胞中的大部分蛋白。
- 未激活时,启动者一般是单体,执行者一般是二聚体。
- 激活启动者有两种方式:外源途径(Extrinsic)和内源途径(Intrinsic,又称线粒体途径)。
- 外源途径:
- 外源途径始于特定信号分子与细胞表面的凋亡受体(Death Receptor)结合,一般信号分子和受体都是同型三聚体。(大部分信号分子都属于肿瘤坏死因子,Tumor Necrosis Factor,TNF)
- 所有受体在细胞内部分都有死亡结构域(Death Domain)。
- TNF中研究最多的成员是TNFα,但它引起凋亡的功能不典型;一般以Fas为典型。
- Fas位于胞毒T细胞和杀手细胞的细胞膜上。
- Fas激活细胞膜上的受体后,受体与FADD蛋白结合,它又和胱天蛋白酶-8(启动者)结合并激活它,形成的复合体称为死亡诱导信号复合体(Death Inducing Signaling Complex,DISC)。
- 人体细胞中,胱天蛋白酶-10也可作为外源途径启动者。
- FLIP蛋白(结构上与胱天蛋白酶-8类似,但没有活性)可抑制外源途径。
- TNFα在激活外源途径的同时,也促进NF-κB复合体的形成,它抑制细胞凋亡,所以TNFα的凋亡作用不典型。
- 除了TNF类分子,还有其它物质可引起凋亡,如蝌蚪尾部的细胞凋亡是由甲状腺激素引起的。
- 内源途径:
- 细胞遭遇DNA损伤或到达发育特定阶段时,自己启动细胞凋亡,此途径称为内源途径。
- 内源途径始于线粒体中的细胞色素c被释放入细胞质,因此内源途径又称线粒体途径。
- 细胞色素c与Apaf1蛋白结合,7个Apaf1聚合成凋亡体(Apoptosome)。
- 凋亡体激活胱天蛋白酶-9(启动者)。
- Bcl2类蛋白负责内源途径的调控,它也是进化上高度保守。
- Bcl2类蛋白有四种结构域,记为BH1,BH2,BH3,BH4。
- 具有BH4结构域的蛋白(Bcl2、BclXL)抑制凋亡,而没有它的(主要是Bax、Bak)促进凋亡。
- Bak始终和线粒体外膜结合,而Bax在细胞质中,它们的激活需要依赖一些只含BH3结构域的蛋白(Bad、Bim、Bid、Puma、Noxa等)。
- Bcl2、BclXL都在线粒体外膜上,通过和没有BH4结构域的蛋白结合而抑制它们发挥功能。
- 只含BH3结构域的蛋白,是Bcl2类蛋白中最大的一类,它们被激活后和Bcl2、BclXL等蛋白相互抑制,从而使Bax和Bak得以发挥功能。
- DNA受损时,细胞产生p53蛋白,它激活Puma和Noxa蛋白的表达,从而引起细胞凋亡。(p53是一种重要的抑癌基因,见癌)
- 有些细胞外的信号也会通过内源途径起作用,起承接作用的是Bid蛋白,它被胱天蛋白酶-8切割后激活。
- IAP类蛋白抑制细胞凋亡。
- IAP类蛋白首先在一些昆虫病毒中被发现,它抑制被感染的细胞凋亡,后来发现大部分动物都有这种蛋白。
- IAP类蛋白的标志结构域是BIR结构域,能与胱天蛋白酶结合并抑制它,有些还能用泛素修饰胱天蛋白酶,使之被降解。
- 对应地,昆虫有抗IAP蛋白(Anti-IAP)。(如果昆虫的IAP或抗IAP有缺陷,则发育不能正常进行,但在哺乳动物中即使完全删去,发育也能正常进行)
- IAP蛋白在细胞质中,抗IAP蛋白在线粒体膜间隙中。
- 有些信号分子抑制细胞凋亡,被称为生存因子(Survival Factor)。
- 很多动物细胞都需要持续不断收到生存因子,才能存活,如神经细胞。
- 大部分生存因子的机制是调控Bcl2类蛋白,有三种机制:
- 促进含BH4结构域的蛋白的合成。
- 将只含BH3结构域的蛋白磷酸化,抑制它。
- 将抗IAP蛋白磷酸化,抑制它。
- 细胞凋亡的启动需要蛋白合成,但凋亡过程中蛋白合成受强烈抑制。
细胞坏死


{kind=link}
图7:细胞凋亡和细胞坏死的区别

{kind=link}
图8A:细胞坏死的形态

{kind=link}
图8B:细胞坏死的形态


{kind=link}
图9:凝固性坏死


{kind=link}
图10:液化性坏死


{kind=link}
图11:干酪样坏死


{kind=link}
图12:纤维蛋白样坏死
- 细胞受到不可逆转的损伤时,启动细胞坏死;细胞坏死一定是细胞在病理条件下的行为。
- 细胞损伤的外部原因通常有:缺氧(Hypoxia)、机械损伤、高温、低温、气压突然改变、辐射、电击、化学损伤(极高浓度的氧气、葡萄糖、氯化钠等都会引起)、寄生物种、免疫反应、遗传病、营养不均衡。
- 细胞损伤的根本原因可归结为:ATP耗尽、线粒体受损、钙离子化学梯度被打破、氧自由基大量积累、细胞膜通透性改变、DNA和蛋白质受损。
- 细胞坏死的形态变化:
- 首先细胞肿胀,即使是可逆损伤也会出现。
- 细胞中出现大量囊泡,可能是内质网破碎。
- 细胞膜形状出现变化,如微绒毛消失。
- 细胞表面显得毛糙,因为糖蛋白颗粒消失。
- 细胞肿胀到一定程度,细胞膜破裂。
- 苏木精-伊红染色显示细胞嗜酸性更强,主要是由于细胞质中RNA被降解,蛋白质被变性。
- DNA降解高度随机,凝胶电泳呈涂抹状条带。
- 细胞核的变化有三种方式:
- 核溶解(Karyolysis):细胞核大小不变,染色质嗜碱性褪去。
- 核固缩(Pyknosis):细胞核缩小,染色质浓缩成一团嗜碱性很强的物质。
- 核碎裂(Karyorrhexis):先发生核固缩,然后细胞核碎成很多块。
- 细胞内容物被释放,引起发炎,来自细胞自身和周围吞噬细胞的溶酶体的酶将这些物质消化,通常需要几个小时。
- 组织坏死的几种形态:
- 凝固性坏死(Coagulative Necrosis):损伤不但使细胞凋亡,还使蛋白酶发生变性,结果坏死细胞释放的物质在很长一段时间内不降解,但它们最终会被渗透入的吞噬细胞清理干净。(一块凝固性坏死区域称为一个梗塞(Infarct))
- 液化性坏死(Liquefactive Necrosis):与凝固性坏死相对,坏死组织很快被酶清理。
- 坏疽性坏死(Gangrenous Necrosis):因缺血引起的坏死,一般呈凝固性坏死,若继发细菌感染则部分出现液化性坏死,称为湿性坏疽(Wet Gangrene)。
- 干酪样坏死(Caseous Necrosis):出现在肺部感染结核杆菌时,中心是一团脆的黄白色物质,周围发炎。
- 纤维蛋白样坏死(Fibrinoid Necrosis):出现在抗原-抗体反应沉淀在动脉壁上时。
- 细胞坏死时蛋白合成不受抑制,只要核糖体不变性就继续合成蛋白。
细胞程序性坏死


{kind=link}
图13:细胞程序性坏死
- 细胞程序性坏死是一些信号分子通过调控细胞的代谢,使细胞受损,然后发生坏死的过程。
- 目前,以TNFα引起的程序性坏死研究最充分,但整个信号通路仍是不确定的。
- TNFα的受体是TNFR-1,它们结合时吸引TRADD、RIPK1(又名RIP1)、TRAF蛋白。
- TRADD蛋白的功能是招募FADD蛋白,即引起细胞凋亡。
- TRAF蛋白能招募cIAP蛋白(一种IAP类蛋白),形成以cIAP为核心的泛素链组装复合体(Linear Ubiquitin Chain Assembly Complex,LUBAC),本质上是一种泛素连接酶。
- LUBAC在RIPK1的赖氨酸残基上组装泛素链,但不会使其被降解。
- RIPK1和TRAF蛋白激活IKK复合体,进而激活NK-κB蛋白,它通过促进合成抑制胱天蛋白酶-8的蛋白来抑制细胞凋亡。
- RIPK1的泛素链抑制细胞坏死。
- 在特定情况下,RIPK1的泛素链被CYLD酶(Cylindromatosis,一种去泛素化酶,因与圆柱瘤病有关而得名)脱去,这会促进细胞坏死。
- 细胞坏死是否发生是由胱天蛋白酶-8的活性决定的,即若NK-κB蛋白成功抑制了胱天蛋白酶-8,则细胞坏死才会发生。
- RIPK1和RIPK3蛋白在被磷酸化后结合,组成坏死体(Necrosome),磷酸化主要是由RIPK1完成的。(RIPK1和RIPK3都是蛋白激酶)
- 坏死体是引起程序性坏死的关键,但它如何引起仍是不清楚的,有人发现坏死体能在细胞中组成淀粉样纤维(就像朊病毒),据此认为坏死体的功能和朊病毒类似。
- 总之,细胞程序性坏死是一种在胱天蛋白酶受抑制的情况下发生的细胞死亡方式。
附录:细胞自噬
- 细胞自噬是细胞把自己的一些细胞器和其它物质送入溶酶体分解的过程。
- 细胞自噬大致有三种:
- 伴侣介导自噬(Chaperone Mediated Autophagy):由分子伴侣帮助将蛋白送入溶酶体。
- 微自噬(Microautophagy):溶酶体的膜内陷将一些物质送入。
- 巨自噬(Macroautophagy):细胞质的一部分被围入一个有双层膜的空间,然后它和溶酶体融合。